

Spatial discontinuity of optomotor-blind expression in the Drosophila wing imaginal disc disrupts epithelial architecture and promotes cell sorting
- PMID: 20178599
- PMCID: PMC2838827
- DOI: 10.1186/1471-213X-10-23
Spatial discontinuity of optomotor-blind expression in the Drosophila wing imaginal disc disrupts epithelial architecture and promotes cell sorting
Abstract
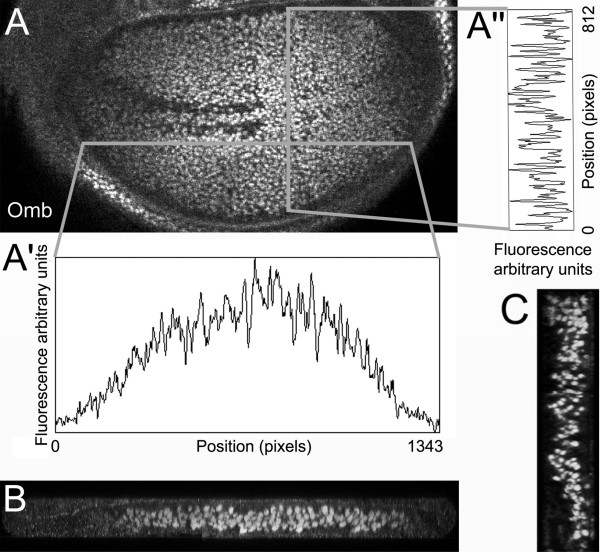
Background: Decapentaplegic (Dpp) is one of the best characterized morphogens, required for dorso-ventral patterning of the Drosophila embryo and for anterior-posterior (A/P) patterning of the wing imaginal disc. In the larval wing pouch, the Dpp target gene optomotor-blind (omb) is generally assumed to be expressed in a step function above a certain threshold of Dpp signaling activity.
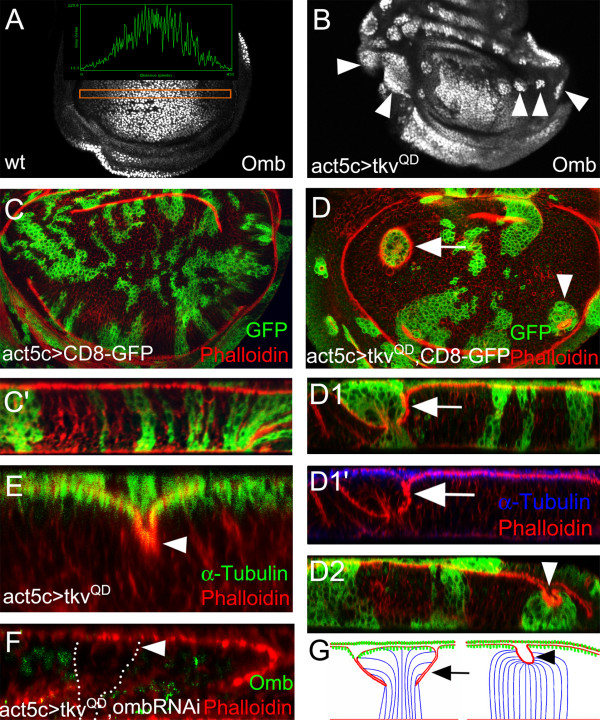
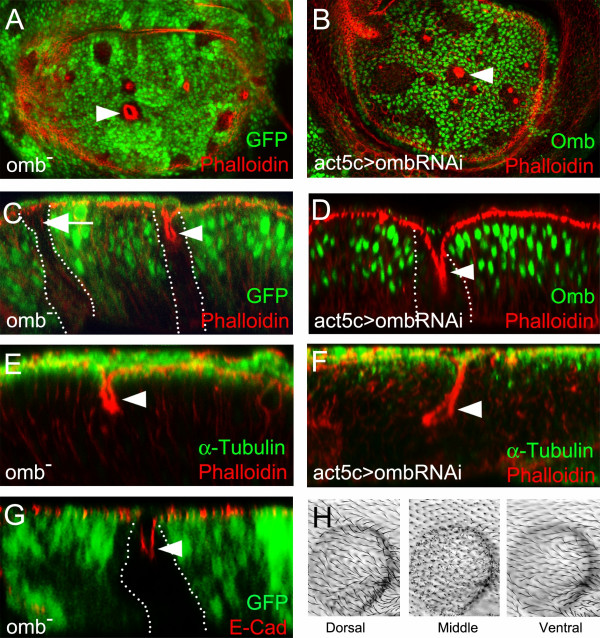
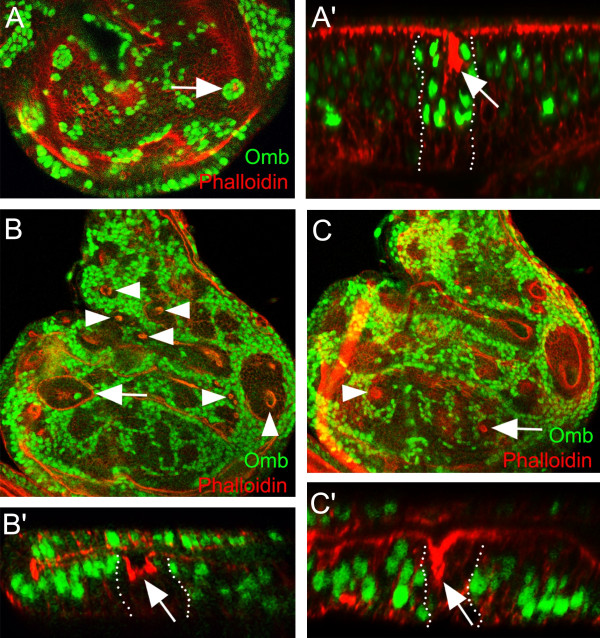
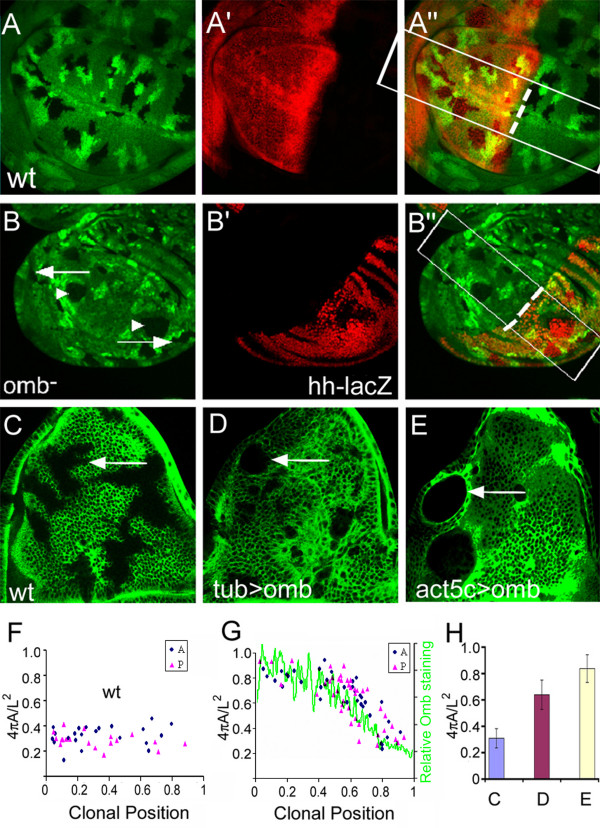
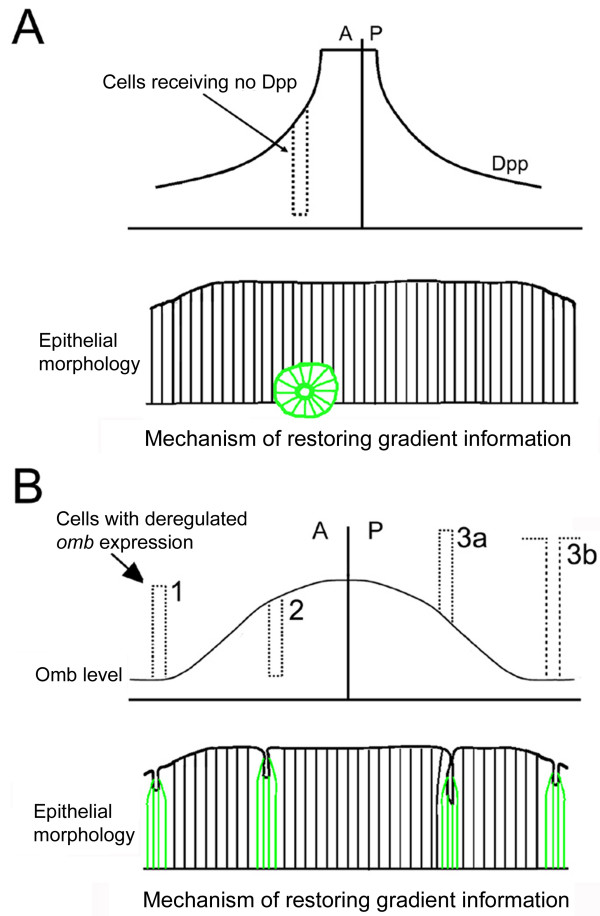
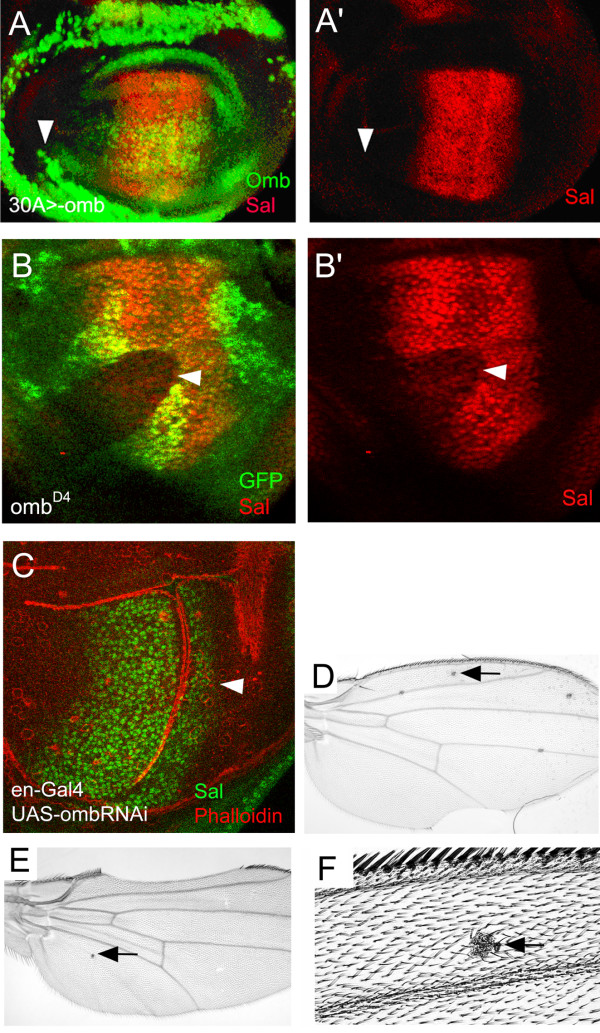
Results: We show that the transcription factor Omb forms, in fact, a symmetrical gradient on both sides of the A/P compartment boundary. Disruptions of the Omb gradient lead to a re-organization of the epithelial cytoskeleton and to a retraction of cells toward the basal membrane suggesting that the Omb gradient is required for correct epithelial morphology. Moreover, by analysing the shape of omb gain- and loss-of-function clones, we find that Omb promotes cell sorting along the A/P axis in a concentration-dependent manner.
Conclusions: Our findings show that Omb distribution in the wing imaginal disc is described by a gradient rather than a step function. Graded Omb expression is necessary for normal cell morphogenesis and cell affinity and sharp spatial discontinuities must be avoided to allow normal wing development.
Figures
Similar articles
-
brinker and optomotor-blind act coordinately to initiate development of the L5 wing vein primordium in Drosophila.Development. 2004 May;131(9):2113-24. doi: 10.1242/dev.01100. Epub 2004 Apr 8. Development. 2004. PMID: 15073155
-
Dpp signaling inhibits proliferation in the Drosophila wing by Omb-dependent regional control of bantam.Development. 2013 Jul;140(14):2917-22. doi: 10.1242/dev.094300. Development. 2013. PMID: 23821035
-
Complementary expression of optomotor-blind and the Iroquois complex promotes fold formation to separate wing notum and hinge territories.Dev Biol. 2016 Aug 1;416(1):225-234. doi: 10.1016/j.ydbio.2016.05.020. Epub 2016 May 19. Dev Biol. 2016. PMID: 27212024
-
T-Box Genes in Drosophila Limb Development.Curr Top Dev Biol. 2017;122:313-354. doi: 10.1016/bs.ctdb.2016.08.003. Epub 2016 Sep 9. Curr Top Dev Biol. 2017. PMID: 28057269 Review.
-
Coordination of patterning and growth by the morphogen DPP.Curr Biol. 2014 Mar 17;24(6):R245-55. doi: 10.1016/j.cub.2014.01.055. Curr Biol. 2014. PMID: 24650915 Review.
Cited by
-
Enhancer trap infidelity in Drosophila optomotor-blind.Fly (Austin). 2013 Apr-Jun;7(2):118-28. doi: 10.4161/fly.23657. Epub 2013 Mar 21. Fly (Austin). 2013. PMID: 23519069 Free PMC article.
-
Fold formation at the compartment boundary of Drosophila wing requires Yki signaling to suppress JNK dependent apoptosis.Sci Rep. 2016 Nov 29;6:38003. doi: 10.1038/srep38003. Sci Rep. 2016. PMID: 27897227 Free PMC article.
-
Bilateral JNK activation is a hallmark of interface surveillance and promotes elimination of aberrant cells.Elife. 2023 Feb 6;12:e80809. doi: 10.7554/eLife.80809. Elife. 2023. PMID: 36744859 Free PMC article.
-
Decapentaplegic and growth control in the developing Drosophila wing.Nature. 2015 Nov 19;527(7578):375-8. doi: 10.1038/nature15730. Epub 2015 Nov 9. Nature. 2015. PMID: 26550824
-
Dpp spreading is required for medial but not for lateral wing disc growth.Nature. 2015 Nov 19;527(7578):317-22. doi: 10.1038/nature15712. Epub 2015 Nov 9. Nature. 2015. PMID: 26550827
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
