

Dicer-independent primal RNAs trigger RNAi and heterochromatin formation
- PMID: 20178743
- PMCID: PMC3020400
- DOI: 10.1016/j.cell.2010.01.019
Dicer-independent primal RNAs trigger RNAi and heterochromatin formation
Abstract
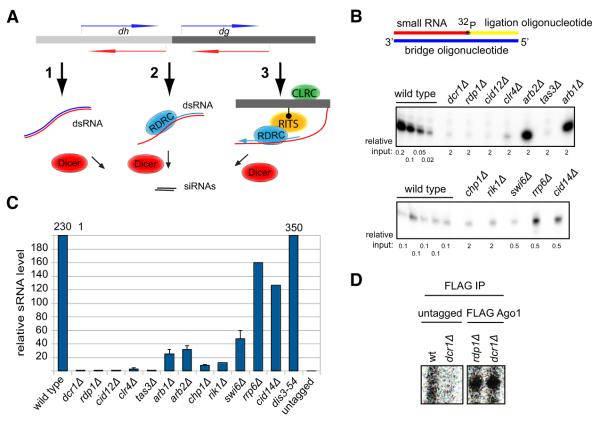
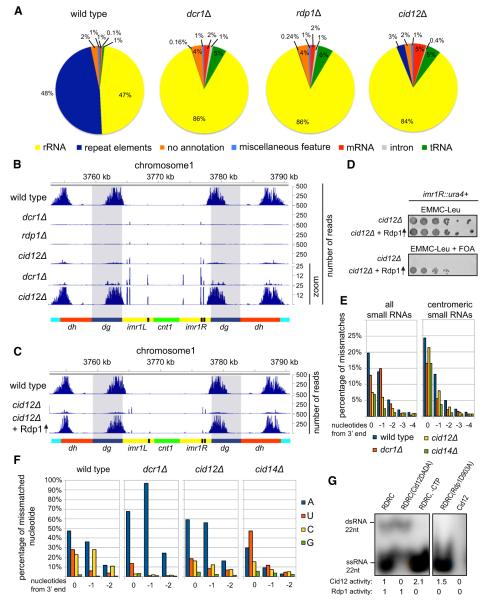
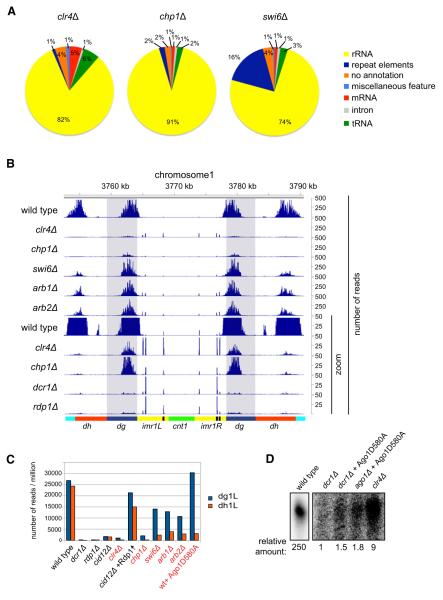
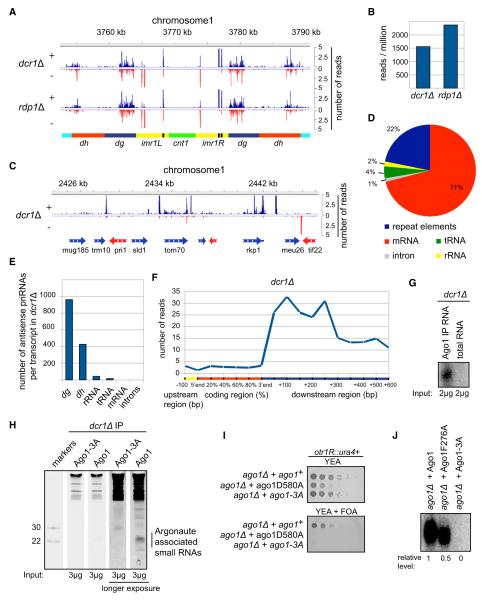
Assembly of fission yeast pericentromeric heterochromatin and generation of small interfering RNAs (siRNAs) from noncoding centromeric transcripts are mutually dependent processes. How this interdependent positive feedback loop is first triggered is a fundamental unanswered question. Here, we show that two distinct Argonaute (Ago1)-dependent pathways mediate small RNA generation. RNA-dependent RNA polymerase complex (RDRC) and Dicer act on specific noncoding RNAs to generate siRNAs by a mechanism that requires the slicer activity of Ago1 but is independent of pre-existing heterochromatin. In the absence of RDRC or Dicer, a distinct class of small RNAs, called primal small RNAs (priRNAs), associates with Ago1. priRNAs are degradation products of abundant transcripts, which bind to Ago1 and target antisense transcripts that result from bidirectional transcription of DNA repeats. Our results suggest that a transcriptome surveillance mechanism based on random association of RNA degradation products with Argonaute triggers siRNA amplification and heterochromatin assembly within DNA repeats.
2010 Elsevier Inc. All rights reserved.
Figures
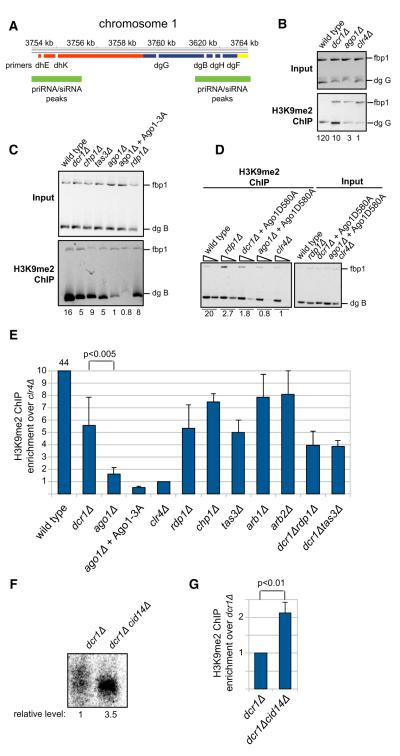
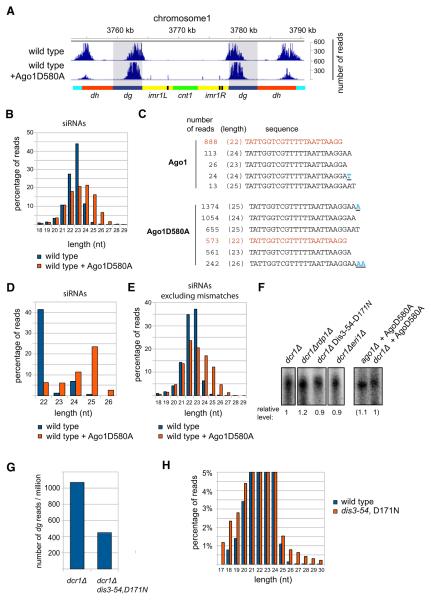
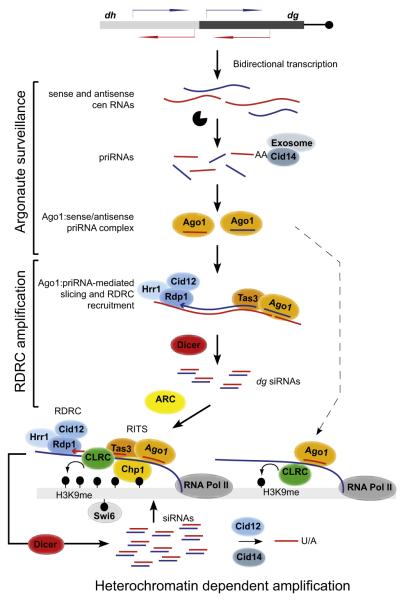
Comment in
-
Primal RNAs: The end of the beginning?Cell. 2010 Feb 19;140(4):452-4. doi: 10.1016/j.cell.2010.02.005. Cell. 2010. PMID: 20178736
References
-
- Aravin AA, Hannon GJ, Brennecke J. The Piwi-piRNA pathway provides an adaptive defense in the transposon arms race. Science. 2007;318:761–764. - PubMed
-
- Baulcombe D. RNA silencing in plants. Nature. 2004;431:356–363. - PubMed
-
- Bühler M, Verdel A, Moazed D. Tethering RITS to a nascent transcript initiates RNAi- and heterochromatin-dependent gene silencing. Cell. 2006;125:873–886. - PubMed
-
- Bühler M, Haas W, Gygi SP, Moazed D. RNAi-dependent and -independent RNA turnover mechanisms contribute to heterochromatic gene silencing. Cell. 2007;129:707–721. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
