

Elevated ATPase activity of KaiC applies a circadian checkpoint on cell division in Synechococcus elongatus
- PMID: 20178745
- PMCID: PMC3031423
- DOI: 10.1016/j.cell.2009.12.042
Elevated ATPase activity of KaiC applies a circadian checkpoint on cell division in Synechococcus elongatus
Abstract
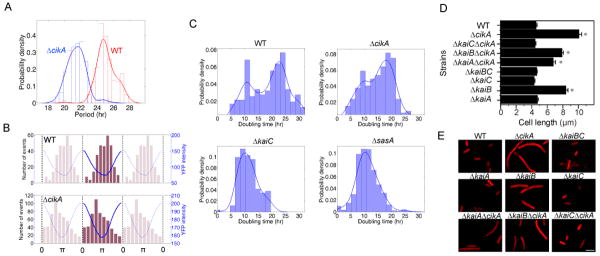
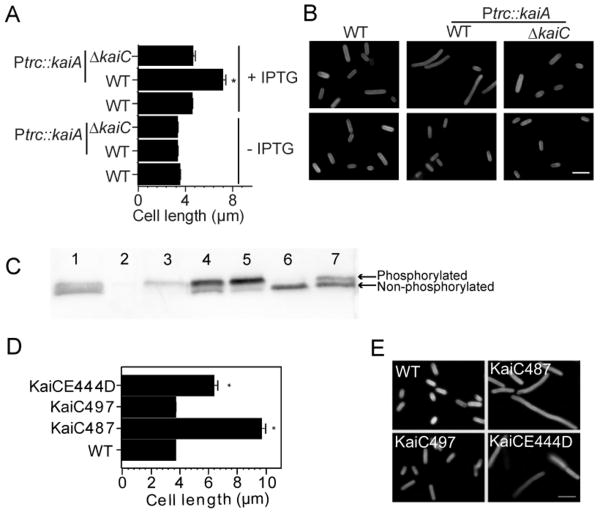
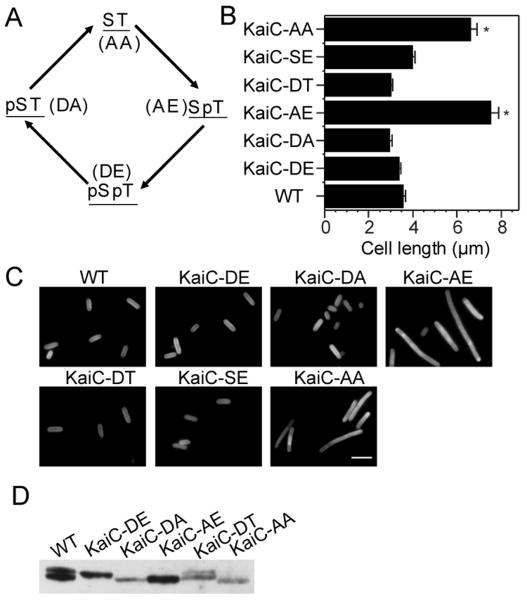
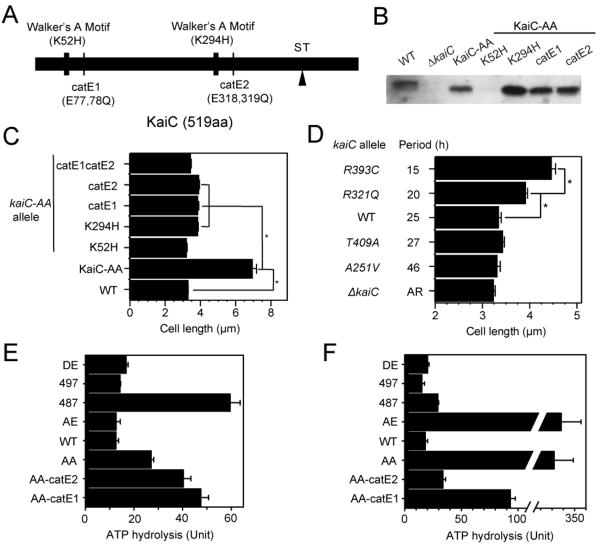
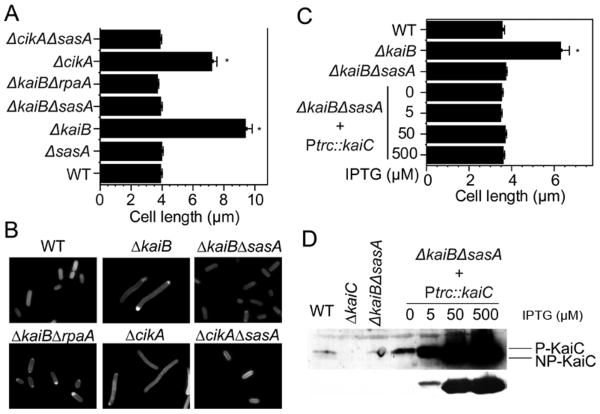
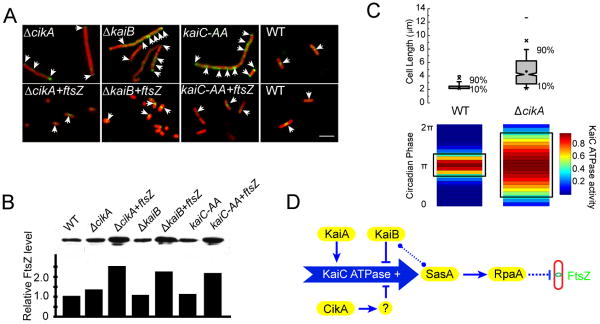
A circadian clock coordinates physiology and behavior in diverse groups of living organisms. Another major cyclic cellular event, the cell cycle, is regulated by the circadian clock in the few cases where linkage of these cycles has been studied. In the cyanobacterium Synechococcus elongatus, the circadian clock gates cell division by an unknown mechanism. Using timelapse microscopy, we confirm the gating of cell division in the wild-type and demonstrate the regulation of cytokinesis by key clock components. Specifically, a state of the oscillator protein KaiC that is associated with elevated ATPase activity closes the gate by acting through a known clock output pathway to inhibit FtsZ ring formation at the division site. An activity that stimulates KaiC phosphorylation independently of the KaiA protein was also uncovered. We propose a model that separates the functions of KaiC ATPase and phosphorylation in cell division gating and other circadian behaviors.
2010 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Circadian cell-cycle progression: Cracking open the gate.Cell. 2010 Feb 19;140(4):458-9. doi: 10.1016/j.cell.2010.02.002. Cell. 2010. PMID: 20178739
References
-
- Chabot JR, Pedraza JM, Luitel P, van Oudenaarden A. Stochastic gene expression out-of-steady-state in the cyanobacterial circadian clock. Nature. 2007;450:1249–1252. - PubMed
-
- Chen Z, Odstrcil EA, Tu BP, McKnight SL. Restriction of DNA replication to the reductive phase of the metabolic cycle protects genome integrity. Science. 2007;316:1916–1919. - PubMed
-
- Clerico EM, Ditty JL, Golden SS. Methods in molecular biology. Vol. 362. Clifton, NJ: 2007. Specialized techniques for site-directed mutagenesis in cyanobacteria; pp. 155–171. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
