

Phosphorylation of the Mycobacterium tuberculosis beta-ketoacyl-acyl carrier protein reductase MabA regulates mycolic acid biosynthesis
- PMID: 20178986
- PMCID: PMC2857119
- DOI: 10.1074/jbc.M110.105189
Phosphorylation of the Mycobacterium tuberculosis beta-ketoacyl-acyl carrier protein reductase MabA regulates mycolic acid biosynthesis
Abstract
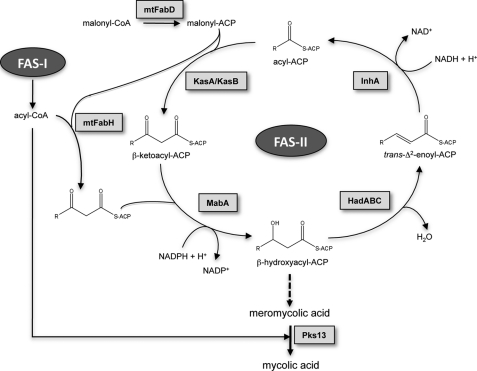
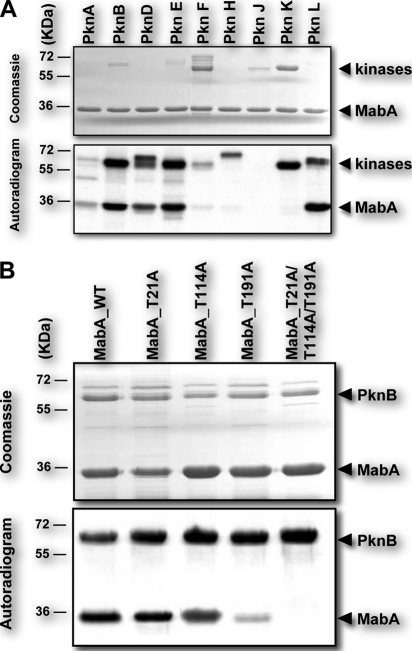
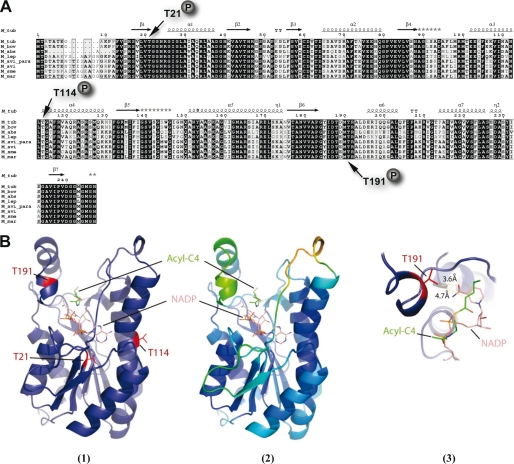
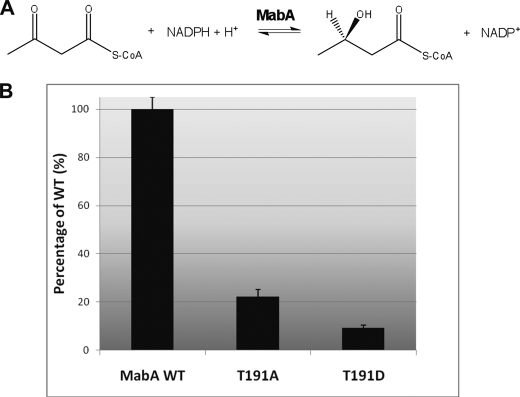
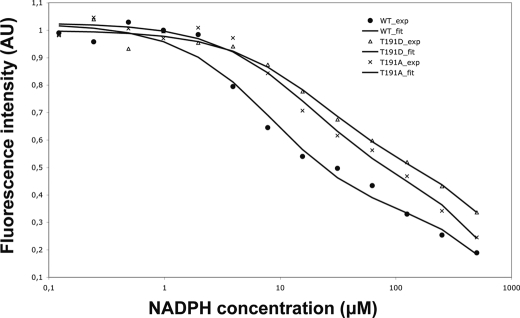
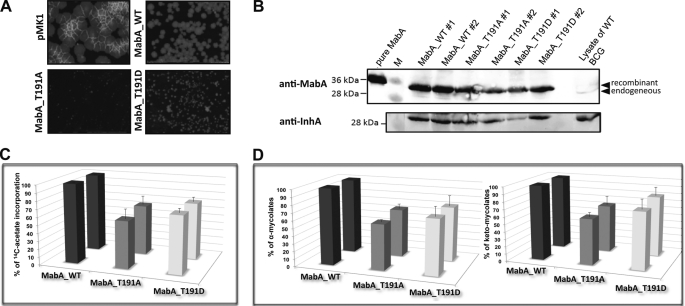
Mycolic acids are key cell wall components for the survival, pathogenicity, and antibiotic resistance of the human tubercle bacillus. Although it was thought that Mycobacterium tuberculosis tightly regulates their production to adapt to prevailing environmental conditions, the molecular mechanisms governing mycolic acid biosynthesis remained extremely obscure. Meromycolic acids, the direct precursors of mycolic acids, are synthesized by a type II fatty acid synthase from acyl carrier protein-bound substrates that are extended iteratively, with a reductive cycle in each round of extension, the second step of which is catalyzed by the essential beta-ketoacyl-acyl carrier protein reductase, MabA. In this study, we investigated whether post-translational modifications of MabA might represent a strategy employed by M. tuberculosis to regulate mycolic acid biosynthesis. Indeed, we show here that MabA was efficiently phosphorylated in vitro by several M. tuberculosis Ser/Thr protein kinases, including PknB, as well as in vivo in mycobacteria. Mass spectrometric analyses using LC-ESI/MS/MS and site-directed mutagenesis identified three phosphothreonines, with Thr(191) being the primary phosphor-acceptor. A MabA_T191D mutant, designed to mimic constitutive phosphorylation, exhibited markedly decreased ketoacyl reductase activity compared with the wild-type protein, as well as impaired binding of the NADPH cofactor, as demonstrated by fluorescence spectroscopy. The hypothesis that phosphorylation of Thr(191) alters the enzymatic activity of MabA, and subsequently mycolic acid biosynthesis, was further supported by the fact that constitutive overexpression of the mabA_T191D allele in Mycobacterium bovis BCG strongly impaired mycobacterial growth. Importantly, conditional expression of the phosphomimetic MabA_T191D led to a significant inhibition of de novo biosynthesis of mycolic acids. This study provides the first information on the molecular mechanism(s) involved in mycolic acid regulation through Ser/Thr protein kinase-dependent phosphorylation of a type II fatty acid synthase enzyme.
Figures
Similar articles
-
The Mycobacterium tuberculosis beta-ketoacyl-acyl carrier protein synthase III activity is inhibited by phosphorylation on a single threonine residue.J Biol Chem. 2009 Mar 6;284(10):6414-24. doi: 10.1074/jbc.M806537200. Epub 2008 Dec 11. J Biol Chem. 2009. PMID: 19074144 Free PMC article.
-
MabA (FabG1), a Mycobacterium tuberculosis protein involved in the long-chain fatty acid elongation system FAS-II.Microbiology (Reading). 2002 Apr;148(Pt 4):951-960. doi: 10.1099/00221287-148-4-951. Microbiology (Reading). 2002. PMID: 11932442
-
Role of Serine140 in the mode of action of Mycobacterium tuberculosis β-ketoacyl-ACP Reductase (MabA).BMC Res Notes. 2012 Sep 25;5:526. doi: 10.1186/1756-0500-5-526. BMC Res Notes. 2012. PMID: 23006410 Free PMC article.
-
The Molecular Genetics of Mycolic Acid Biosynthesis.Microbiol Spectr. 2014 Aug;2(4):MGM2-0003-2013. doi: 10.1128/microbiolspec.MGM2-0003-2013. Microbiol Spectr. 2014. PMID: 26104214 Review.
-
Kinase Targets for Mycolic Acid Biosynthesis in Mycobacterium tuberculosis.Curr Mol Pharmacol. 2019;12(1):27-49. doi: 10.2174/1874467211666181025141114. Curr Mol Pharmacol. 2019. PMID: 30360731 Review.
Cited by
-
Eukaryote-like serine/threonine kinases and phosphatases in bacteria.Microbiol Mol Biol Rev. 2011 Mar;75(1):192-212. doi: 10.1128/MMBR.00042-10. Microbiol Mol Biol Rev. 2011. PMID: 21372323 Free PMC article. Review.
-
Multisystem Analysis of Mycobacterium tuberculosis Reveals Kinase-Dependent Remodeling of the Pathogen-Environment Interface.mBio. 2018 Mar 6;9(2):e02333-17. doi: 10.1128/mBio.02333-17. mBio. 2018. PMID: 29511081 Free PMC article.
-
Phosphorylation of KasB regulates virulence and acid-fastness in Mycobacterium tuberculosis.PLoS Pathog. 2014 May 8;10(5):e1004115. doi: 10.1371/journal.ppat.1004115. eCollection 2014 May. PLoS Pathog. 2014. PMID: 24809459 Free PMC article.
-
Epigenetic Phosphorylation Control of Mycobacterium tuberculosis Infection and Persistence.Microbiol Spectr. 2017 Mar;5(2):10.1128/microbiolspec.tbtb2-0005-2015. doi: 10.1128/microbiolspec.TBTB2-0005-2015. Microbiol Spectr. 2017. PMID: 28281439 Free PMC article. Review.
-
Hanks-Type Serine/Threonine Protein Kinases and Phosphatases in Bacteria: Roles in Signaling and Adaptation to Various Environments.Int J Mol Sci. 2018 Sep 21;19(10):2872. doi: 10.3390/ijms19102872. Int J Mol Sci. 2018. PMID: 30248937 Free PMC article. Review.
References
-
- Frieden T. R., Sterling T. R., Munsiff S. S., Watt C. J., Dye C. (2003) Lancet 362, 887–899 - PubMed
-
- Flynn J. L., Chan J. (2003) Curr. Opin. Immunol. 15, 450–455 - PubMed
-
- Daffé M., Draper P. (1998) Adv. Microb. Physiol. 39, 131–203 - PubMed
-
- Kremer L., Baulard A. R., Besra G. S. (2000) in Molecular Genetics of Mycobacteria (Hatfull G. F., Jacobs W. R., Jr. eds) pp. 173–190, ASM Press, Washington, DC
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
