

DNA-PKcs controls an endosomal signaling pathway for a proinflammatory response by natural killer cells
- PMID: 20179272
- PMCID: PMC4793721
- DOI: 10.1126/scisignal.2000467
DNA-PKcs controls an endosomal signaling pathway for a proinflammatory response by natural killer cells
Abstract
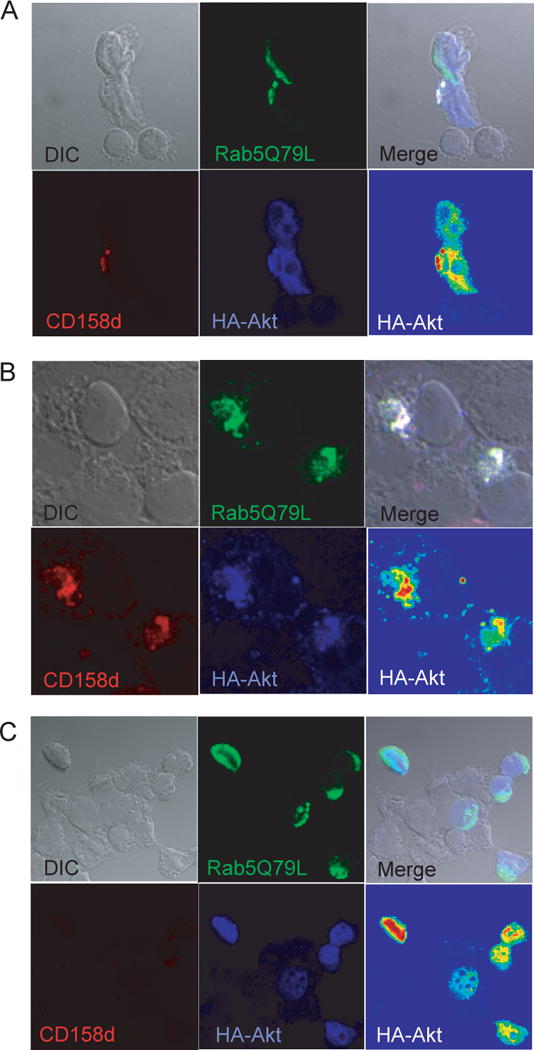
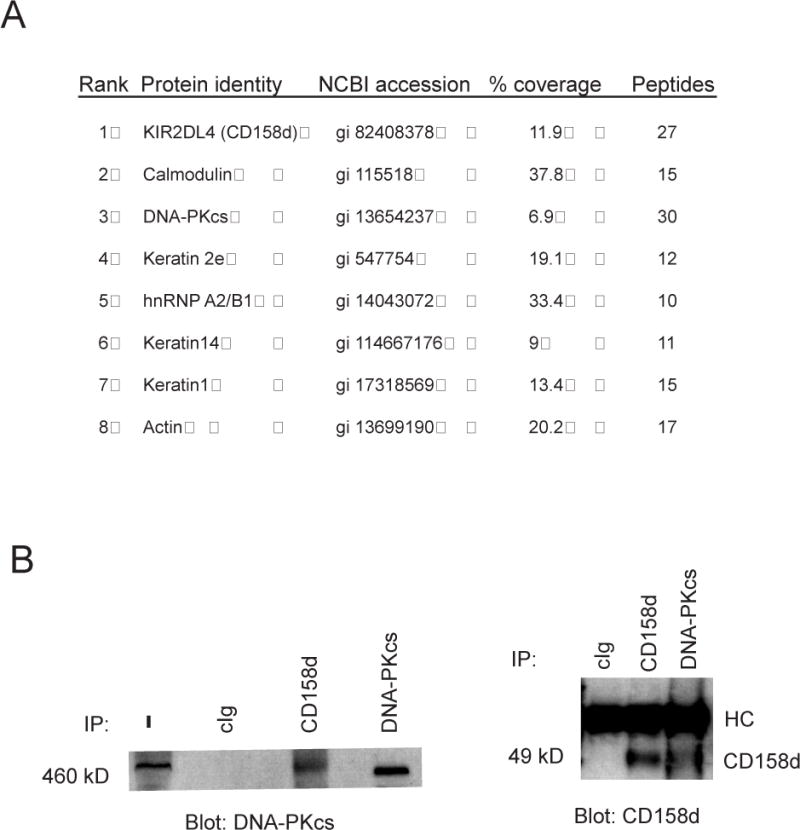
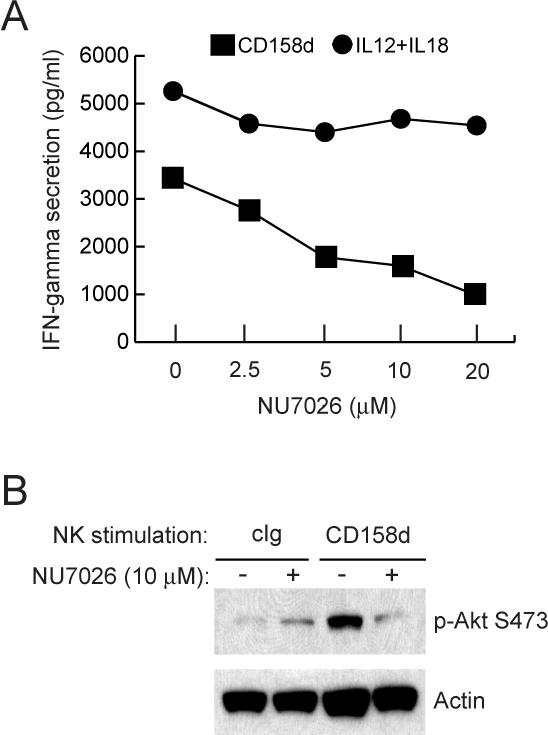
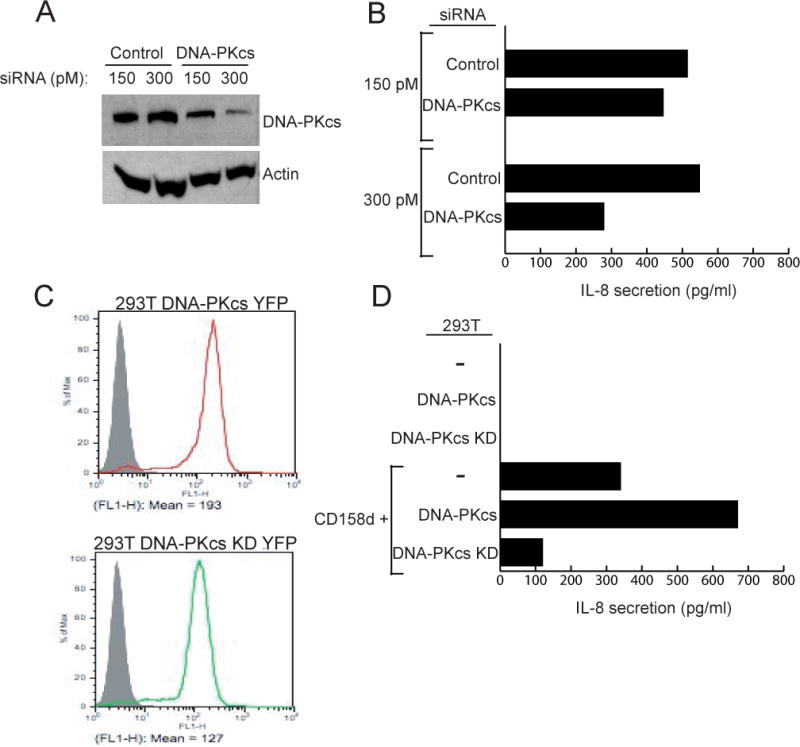
Endosomes are emerging as specialized signaling compartments that endow receptors with distinct signaling properties. The diversity of endosomal signaling pathways and their contribution to various biological responses is still unclear. CD158d, which is also known as the killer cell immunoglobulin-like receptor (KIR) 2DL4 (KIR2DL4), is an endosome-resident receptor in natural killer (NK) cells that stimulates the release of a unique set of proinflammatory and proangiogenic mediators in response to soluble human leukocyte antigen G (HLA-G). Here, we identified the CD158d signaling cascade. In response to soluble agonist antibody or soluble HLA-G, signaling by CD158d was dependent on the activation of nuclear factor kappaB (NF-kappaB) and the serine-threonine kinase Akt. CD158d associated with the catalytic subunit of DNA-dependent protein kinase (DNA-PKcs), promoted the recruitment of Akt to endosomes, and stimulated the DNA-PKcs-dependent phosphorylation of Akt. The sequential requirement for DNA-PKcs, Akt, and NF-kappaB in signaling by CD158d delineates a previously uncharacterized endosomal signaling pathway for a proinflammatory response in NK cells.
Figures
References
-
- Miaczynska M, Pelkmans L, Zerial M. Not just a sink: endosomes in control of signal transduction. Curr Opin Cell Biol. 2004;16:400–406. - PubMed
-
- Polo S, Di Fiore PP. Endocytosis conducts the cell signaling orchestra. Cell. 2006;124:897–900. - PubMed
-
- Hoeller D, Volarevic S, Dikic I. Compartmentalization of growth factor receptor signalling. Curr Opin Cell Biol. 2005;17:107–111. - PubMed
-
- Sorkin A, Von Zastrow M. Signal transduction and endocytosis: close encounters of many kinds. Nat Rev Mol Cell Biol. 2002;3:600–614. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
