

Review
doi: 10.1038/nrc2806.
Eph receptors and ephrins in cancer: bidirectional signalling and beyond
Affiliations
- PMID: 20179713
- PMCID: PMC2921274
- DOI: 10.1038/nrc2806
Item in Clipboard
Review
Eph receptors and ephrins in cancer: bidirectional signalling and beyond
Nat Rev Cancer.
2010 Mar.
Abstract
The Eph receptor tyrosine kinases and their ephrin ligands have intriguing expression patterns in cancer cells and tumour blood vessels, which suggest important roles for their bidirectional signals in many aspects of cancer development and progression. Eph gene mutations probably also contribute to cancer pathogenesis. Eph receptors and ephrins have been shown to affect the growth, migration and invasion of cancer cells in culture as well as tumour growth, invasiveness, angiogenesis and metastasis in vivo. However, Eph signalling activities in cancer seem to be complex, and are characterized by puzzling dichotomies. Nevertheless, the Eph receptors are promising new therapeutic targets in cancer.
Figures
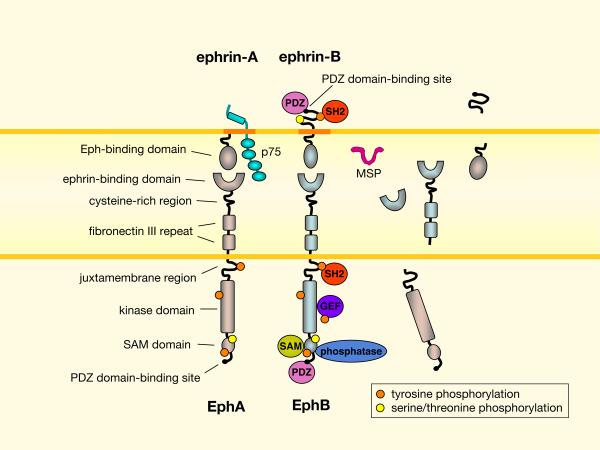
Domain structures of Eph receptors and ephrins are shown. Additionally, alternative splicing or proteolysis can generate extracellular and intracellular domain fragments of Eph receptors and ephrins of both A and B classes. The major sperm protein (MSP) domain from VAMP (vesicle-associated membrane protein)-associated protein (VAP) proteins is another Eph ligand that can compete with ephrins for binding. Eph receptor “forward” signaling involves ephrin-induced clustering, autophosphorylation, and association with signaling effectors containing SH2, PDZ, SAM and other protein interaction domains,. Some signaling proteins, such as certain guanine nucleotide exchange factors (GEFs) for Rho family GTPases, can constitutively associated with Eph receptors. The activities of some effectors are modified by activated Eph receptors, for example through phosphorylation. Phosphotyrosine phosphatases bind Eph receptors and ephrins to dampen or terminate their activity through dephosphorylation. Eph receptors are also phosphorylated on serine/threonine residues, which can have dramatic functional consequences. The transmembrane ephrin-Bs mediate “reverse” signals, which involve Src-dependent tyrosine phosphorylation of their cytoplasmic segment and association with SH2 and PDZ domain-containing proteins,. EphB binding can also affect ephrin-B function by inducing serine phosphorylation, as shown in neurons. The glycosylphosphatidylinositol (GPI)-linked ephrin-As also mediate reverse signals, through poorly understood signaling interactions that may occur in lipid rafts (dark orange). In neurons, ephrin-As can use the p75 nerve growth factor receptor as a signaling partner to activate the FYN Src family kinase. Most domain names are illustrated on EphA/ephrin-A and signaling interactions are illustrated on EphB/ephrin-B, but each applies to the other.
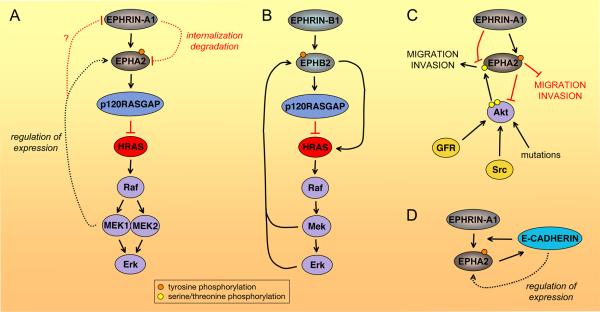
(A) EPHA2/HRAS-Erk negative feedback loop. Activation of the HRAS-Erk pathway increases EPHA2 expression through MEK1 and decreases EPHRIN-A1 expression, although it is not known if this also occurs through MEK1,,. In turn, ephrin-dependent EPHA2 activation inhibits HRAS-Erk signaling and also downregulates EPHA2 levels by causing receptor internalization and degradation. (B) Positive and negative EPHB2/MAP kinase feedback loops. In a positive feedback loop, ephrin-B-dependent EPHB2 activation stimulates the HRAS-Erk pathway, and the increase in Mek and/or Erk activity in turn enables enhanced responsiveness of EPHB2 to ephrin-B stimulation through unknown mechanisms. However, in a different cellular context EPHB2 can also inhibit the HRAS-Erk pathway,, which may in turn reduce EPHB2 activation by ephrin. Although not shown in the figure, EPHRIN-B1 stimulation can also downregulate EphB2 levels by causing internalization/degradation. (C) EPHA2/Akt negative feedback loop. The Akt kinase (activated by growth factor receptors (GFR), Src family kinases or mutations in upstream proteins or Akt itself) phosphorylates serine 897 in the carboxy-terminal tail of EPHA2 leading to increased EPHA2-dependent cell migration and invasion. In turn, EPHRIN-A1-induced EPHA2 signaling inactivates Akt by causing its dephosphorylation at T308 and S473, thus decreasing EPHA2 phosphorylation at serine 897 and, consequently, cell migration and invasion. Other pathways downstream of EPHA2 can also inhibit migration and invasion. (D) EPHA2/E-CADHERIN positive feedback loop. E-CADHERIN expression increases EPHA2 expression, surface localization, interaction with EPHRIN-A1, and thus forward signaling, (Box 1). In turn, EPHA2 signaling enhances E-CADHERIN-mediated adhesion. Dotted lines indicate regulation of protein levels rather than activity.
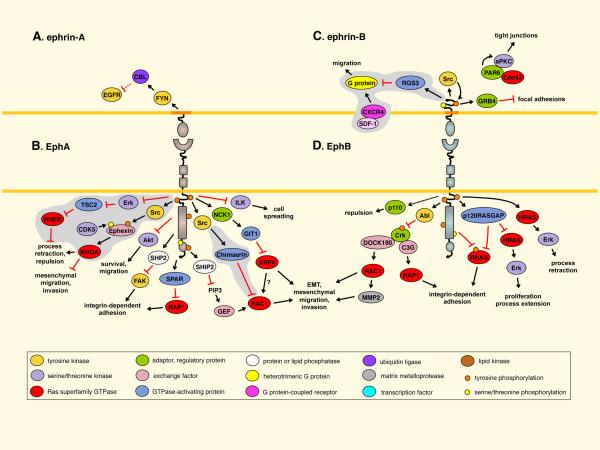
A. EPHRIN-A5 reverse signaling downregulates epidermal growth factor receptor (EGFR) levels in glioma cells. B. EphA receptors activate tuberous sclerosis complex 2 (TSC2) in neurons to inactivate RHEB. EphA activation of RHOA involves Ephexin family exchange factors and other pathways (Fig. 4),. EPHA2 inhibits Akt, and inactivates focal adhesion kinase (FAK) through the SHP2 phosphatase. EPHA4 inhibits RAP1 through spine-associated RAPGAP (SPAR),. Recruitment of the lipid phosphatase SHIP2 by EPHA2 inhibits RAC1 and EPHA2 internalization. EPHA4 inhibits RAC1 through Chimaerins,. EPHA2-mediated inhibition of ADP-ribosylation factor 6 (ARF6) in epithelial cells inhibits epithelial-to-mesenchymal transition (EMT). EphA1 inhibits integrin-linked kinase (ILK). C. EPHRIN-B1 disrupts focal adhesions through GRB4. Phosphorylation inhibits EPHRIN-B1 binding to PAR6, allowing PAR6 to bind GTP-bound CDC42 and activate atypical PKC (aPKC). Ephrin-Bs also inhibit signaling by the CXCR4 G protein-coupled chemokine receptor. D. EphB signaling increases expression of the p110 subunit of PI3 kinase. EphB receptors (and EPHA2) activate Abl, which ultimately inhibits RAP1 and RAC1,,. EPHB2 inactivates RRAS through phosphorylation. EPHB2 (and EPHA2) activates p120RASGAP to inhibit the HRAS and RRAS,. Finally, EPHB2 can activate Erk. Some pathways are assembled from different sources, so the complete pathways are hypothetical. Pathways identified in neurons, and predicted to have tumor suppressing activity, are on gray background. Most other pathways were identified in cultured cells and their significance in cancer also remains to be proven. Dotted lines indicate regulation of expression levels. For more details see references 1,–,,. CDK5, cyclin-dependent kinase 5; GIT1, G protein-coupled receptor kinase-interacting ARFGAP 1; MMP2, matrix metalloproteinase 2; RAPGEF1, Rap guanine nucleotide exchange factor 1; RGS3, regulator of G-protein signaling 3; SDF-1, stromal cell-derived factor-1.
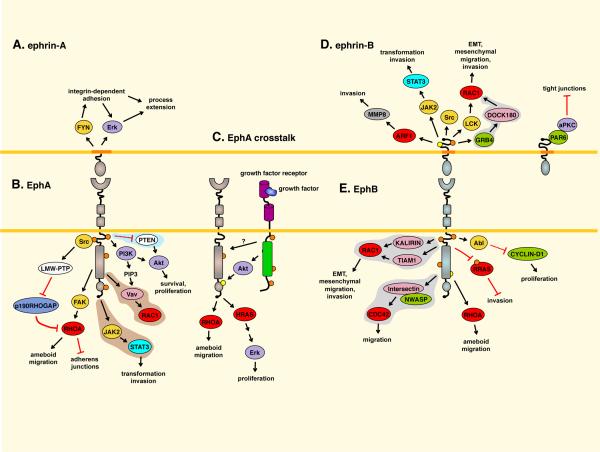
A. EPHRIN-A5 reverse signaling promotes activation of FYN, β1-integrins, and Erk in fibroblasts. B. Low-molecular-weight phosphotyrosine phosphatase (LMW-PTP) is activated by Src and dephosphorylates and inactivates p190RHOGAP. This increases RHOA activity to destabilize adherens junctions in EPHA2-overexpressing epithelial cells. EPHA2 (and EPHB2) activate RHOA through focal adhesion kinase (FAK),. EPHA4 activates signal transducer and activator of transcription 3 (STAT3). A pathway involving EPHA2, PI3 kinase and Vav family exchange factors for RAC1 operates in endothelial cells,. Activation of EPHA2 activates Akt in pancreatic cancer cells. The C. elegans Eph receptor inhibits PTEN expression. C. EPHA2-ERBB2 crosstalk activates the HRAS-Erk pathway and RHOA in a mouse mammary tumor model, enhancing tumor growth and in vitro cell proliferation and migration,. Akt, activated by ERBB2 or other pathways, phosphorylates EPHA2. D. Ephrin-B reverse signaling affects pathways that promote invasiveness, including matrix metalloproteinase 8 (MMP8) secretion and activation of STAT3, Src and RAC1113114. In contrast, non-phosphorylated EPHRIN-B1 can bind PAR6 to inhibit atypical protein kinase C (aPKC). E. EphB forward signaling activates various RAC1 and CDC42 exchange factors,,, which could promote cancer cell migration and invasion. EphB4 activates RHOA. EPHB2-mediated RRAS tyrosine phosphorylation increases glioma cell invasiveness. EPHB2-mediated ABL activation increases CYCLIN-D1 levels. Pathways identified in neurons, endothelial/muscle cells, or C. elegans that are predicted to have tumor promoting activity are on gray, light-brown or light blue background, respectively. Most other pathways were identified in cultured cells and their significance in cancer remains to be proven. For more details see references 1,–,. ARF1, ADP-ribosylation factor 1; EMT, epithelial-to-mesenchymal transition; JAK2, Janus kinase 2.
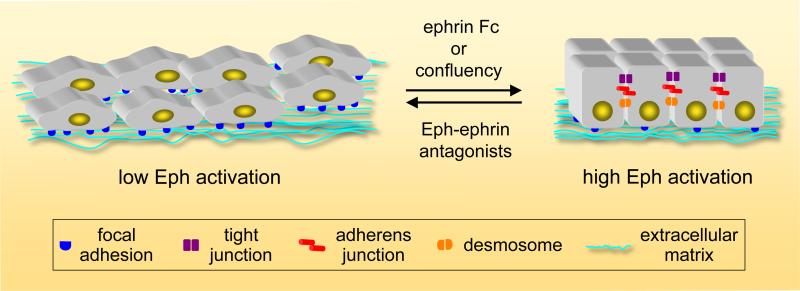
References
-
- Pasquale EB. Eph-ephrin bidirectional signaling in physiology and disease. Cell. 2008;133:38–52. - PubMed
-
- Hirai H, Maru Y, Hagiwara K, Nishida J, Takaku F. A novel putative tyrosine kinase receptor encoded by the eph gene. Science. 1987;238:1717–1720. - PubMed
-
- Maru Y, Hirai H, Takaku F. Overexpression confers an oncogenic potential upon the eph gene. Oncogene. 1990;5:445–447. - PubMed
-
- Bartley TD, et al. B61 is a ligand for the ECK receptor protein-tyrosine kinase. Nature. 1994;368:558–560. - PubMed
-
- Pasquale EB. Eph receptor signalling casts a wide net on cell behaviour. Nat Rev Mol Cell Biol. 2005;6:462–475. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
