

Abnormal post-translational and extracellular processing of brevican in plaque-bearing mice over-expressing APPsw
- PMID: 20180882
- PMCID: PMC2855738
- DOI: 10.1111/j.1471-4159.2010.06647.x
Abnormal post-translational and extracellular processing of brevican in plaque-bearing mice over-expressing APPsw
Abstract
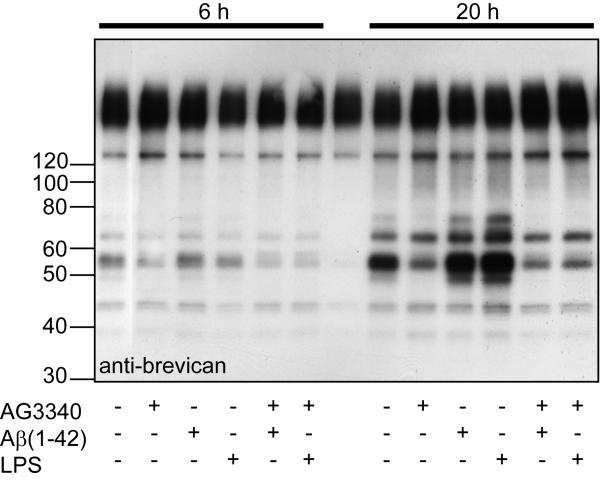
Aggregation of amyloid-beta (Abeta) in the forebrain of Alzheimer's disease (AD) subjects may disturb the molecular organization of the extracellular microenvironment that modulates neural and synaptic plasticity. Proteoglycans are major components of this extracellular environment. To test the hypothesis that Abeta, or another amyloid precursor protein (APP) dependent mechanism modifies the accumulation and/or turnover of extracellular proteoglycans, we examined whether the expression and processing of brevican, an abundant extracellular, chondroitin sulfate (CS)-bearing proteoglycan, were altered in brains of Abeta-depositing transgenic mice (APPsw - APP gene bearing the Swedish mutation) as a model of AD. The molecular size of CS chains attached to brevican was smaller in hippocampal tissue from APPsw mice bearing Abeta deposits compared to non-transgenic mice, likely because of changes in the CS chains. Also, the abundance of the major proteolytic fragment of brevican was markedly diminished in extracts from several telencephalic regions of APPsw mice compared to non-transgenic mice, yet these immunoreactive fragments appeared to accumulate adjacent to the plaque edge. These results suggest that Abeta or APP exert inhibitory effects on proteolytic cleavage mechanisms responsible for synthesis and turnover of proteoglycans. As proteoglycans stabilize synaptic structure and inhibit molecular plasticity, defective brevican processing observed in Abeta-bearing mice and potentially end-stage human AD, may contribute to deficient neural plasticity.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
