

doi: 10.1098/rsbl.2009.0995.
Epub 2010 Feb 24.
Organization and physiology of posterior lateral line afferent neurons in larval zebrafish
Affiliations
- PMID: 20181553
- PMCID: PMC2880066
- DOI: 10.1098/rsbl.2009.0995
Item in Clipboard
Organization and physiology of posterior lateral line afferent neurons in larval zebrafish
Biol Lett.
.
Abstract
The lateral line system of larval zebrafish can translate hydrodynamic signals from the environment to guide body movements. Here, I demonstrate a spatial relationship between the organization of afferent neurons in the lateral line ganglion and the innervation of neuromasts along the body. I developed a whole cell patch clamp recording technique to show that afferents innervate multiple direction-sensitive neuromasts, which are sensitive to low fluid velocities. This work lays the foundation to integrate sensory neuroscience and the hydrodynamics of locomotion in a model genetic system.
Figures
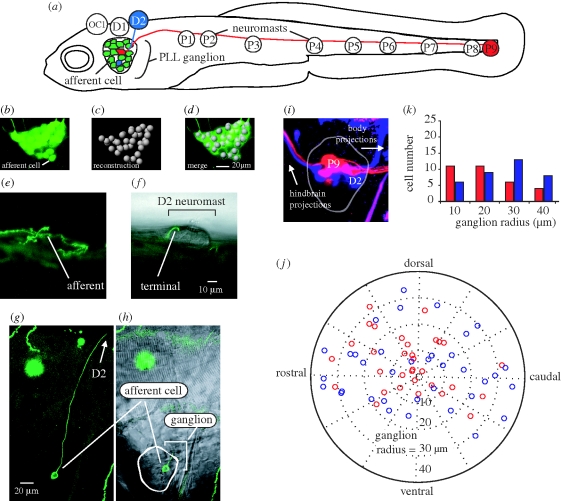
(a) Afferent neurons in the posterior lateral line ganglion of 5 dpf larval zebrafish illustrating connections with D2 and P9 neuromasts (not drawn to scale). Bipolar afferents make projections that terminate on one or more neuromasts labelled OC1, D1-2 and P1-9 (Raible & Kruse 2000; Ledent 2002) as well as to the hindbrain (not shown). (b–d) Afferent cell bodies in a ganglion can be recognized and reconstructed using HUC-GFP transgenic fish. (e,g) DNA injection at the one-celled embryo stage reveals expression of a single afferent innervating the D2 neuromast. (f,h) The position of the afferent in the ganglion and its bulged termini on the neuromast is revealed by merged confocal and Nomarski microscopy images. (i,j) Backfilling dyes into D2 (blue) and P9 (red) neuromasts results in afferent cell body labelling and reveals their relative positions in the ganglion. (k) Graph showing the number of backfilled afferent neurons located in 10 µm concentric ring regions from the centre of the ganglion (n = 14 ganglia). More P9 afferent neurons reside towards the centre of the ganglion while more D2 cells reside in the outer rings of the ganglion.
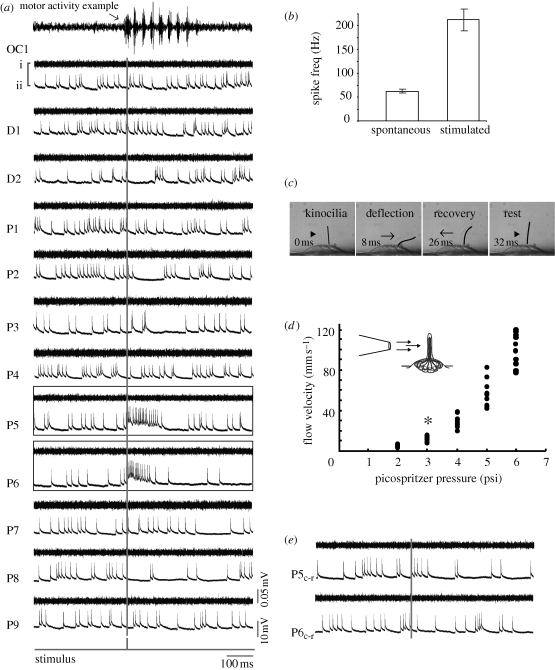
Deflecting a neuromast with a water jet increases firing rate in afferent neurons. (a) Ventral motor root (i) and whole-cell patch clamp recordings of an afferent neuron (ii) while a jet is directed rostrocaudally at each neuromast (rostral is to the left). The stimulus trigger is shown as a grey vertical line. No motor activity occurs during neuromast stimulation. For comparison, an example of swimming activity is shown in the top-most trace. Afferent neurons display spontaneous firing activity even in the absence of a jet. Out of all neuromasts stimulated, only P5 and P6 show an increased firing rate response. (b) All afferents increase their firing rate when the neuromasts that they innervate are stimulated compared to their spontaneous firing rate in the absence of a jet. (c) Deflection of hair cell kinocilia in the D2 neuromast by a jet. The caudal-most kinocilium is traced in black for clarity. (d) Relationship between pressure and ejected water velocity for the stimulus pipette, showing the minimum velocity to elicit afferent firing (asterisk). (e) When the jet is reversed and directed caudorostrally (Pc-r) the afferent neuron does not increase its firing rate. Motor root recordings confirm that no spontaneous firing activity occurs during the experiment, which would decrease neuromast sensitivity due to efferent activity.
References
-
- Alexandre D., Ghysen A.1999Somatotopy of the lateral line projection in larval zebrafish. Proc. Natl Acad. Sci. USA 96, 7558–7562 (doi:10.1073/pnas.96.13.7558) - DOI - PMC - PubMed
-
- Catton K. B., Webster D. R., Brown J., Yen J.2007Quantitative analysis of tethered and free-swimming copepodid flow fields. J. Exp. Biol. 210, 299–310 (doi:10.1242/jeb.02633) - DOI - PubMed
-
- Chagnaud B. P., Bleckmann H., Engelmann J.2006Neural responses of goldfish lateral line afferents to vortex motions. J. Exp. Biol. 209, 327–342 (doi:10.1242/jeb.01982) - DOI - PubMed
-
- Coombs S., Conley R. A.1997Dipole source localization by mottled sculpin. I. Approach strategies. J. Comp. Physiol. A 180, 387–399 (doi:10.1007/s003590050057) - DOI - PubMed
-
- Coombs S., Montgomery J. C.1992Fibers innervating different parts of the lateral line system of the Antarctic fish, Trematomus bernacchii, have similar neural responses despite large variations in peripheral morphology. Brain Behav. Evol. 40, 217–233 (doi:10.1159/000113914) - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases