

The coxsackievirus-adenovirus receptor reveals complex homophilic and heterophilic interactions on neural cells
- PMID: 20181587
- PMCID: PMC6633923
- DOI: 10.1523/JNEUROSCI.5725-09.2010
The coxsackievirus-adenovirus receptor reveals complex homophilic and heterophilic interactions on neural cells
Abstract
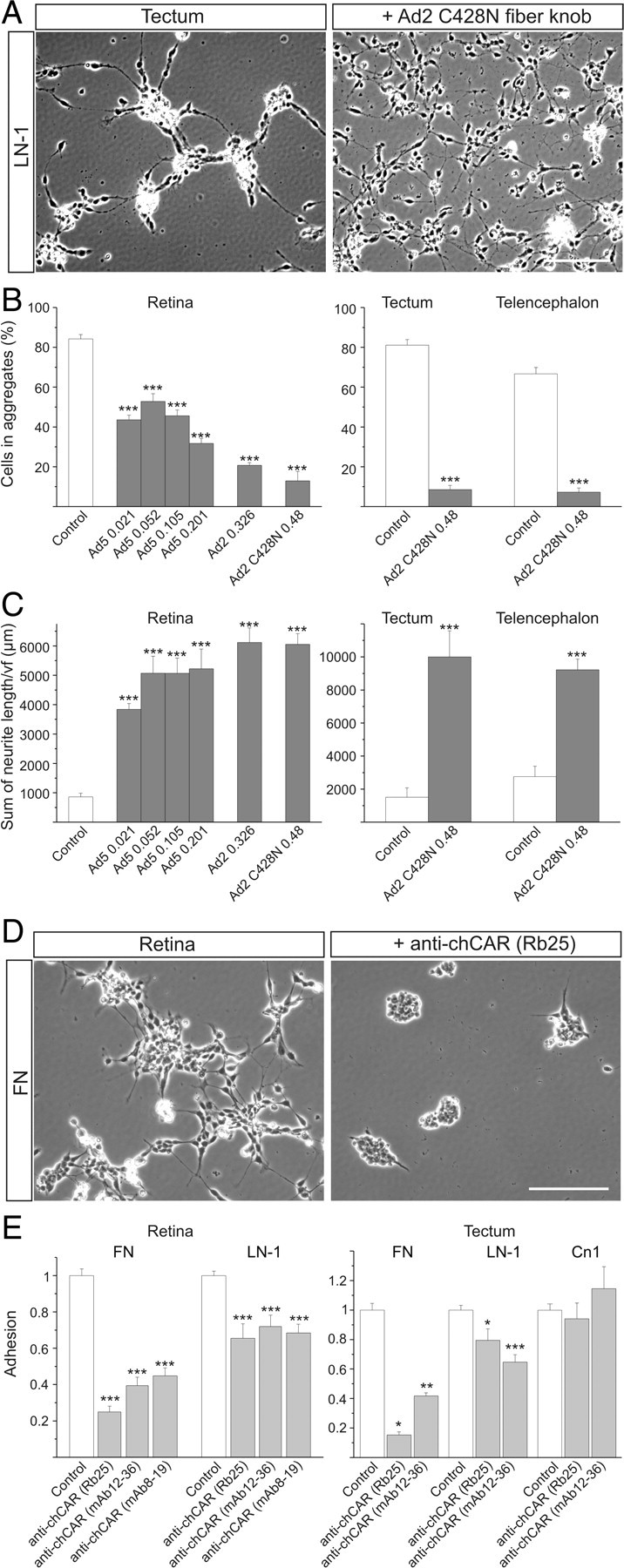
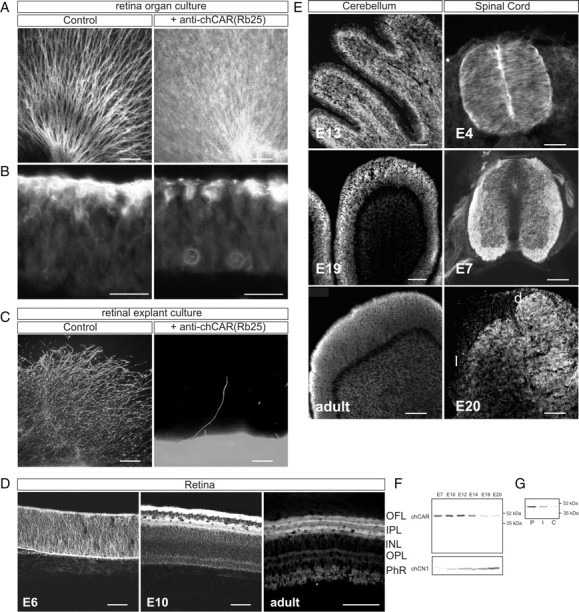
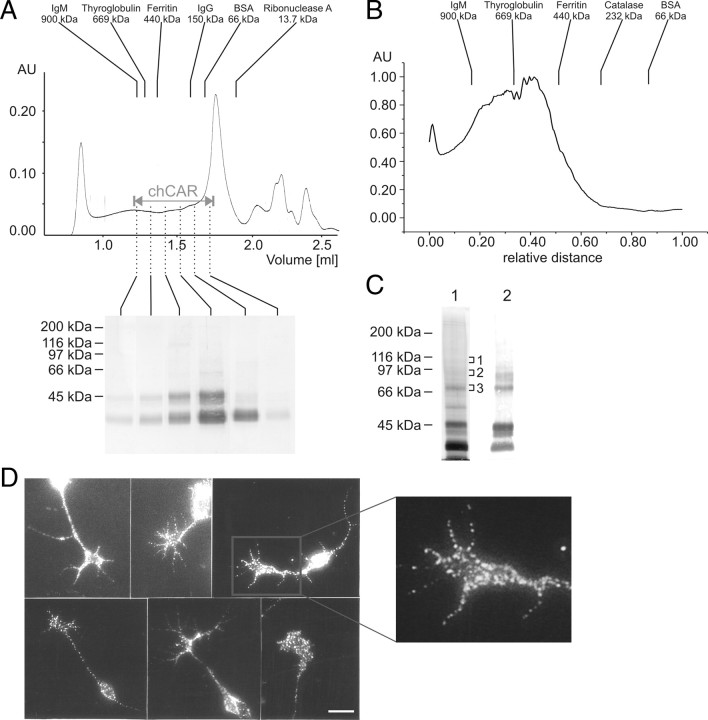
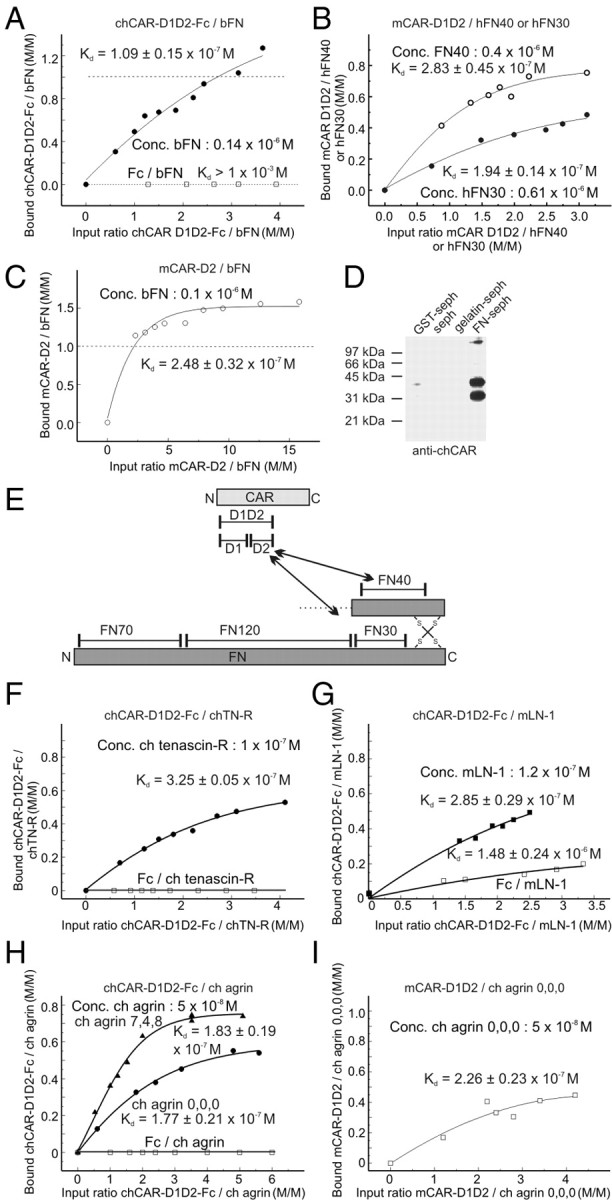
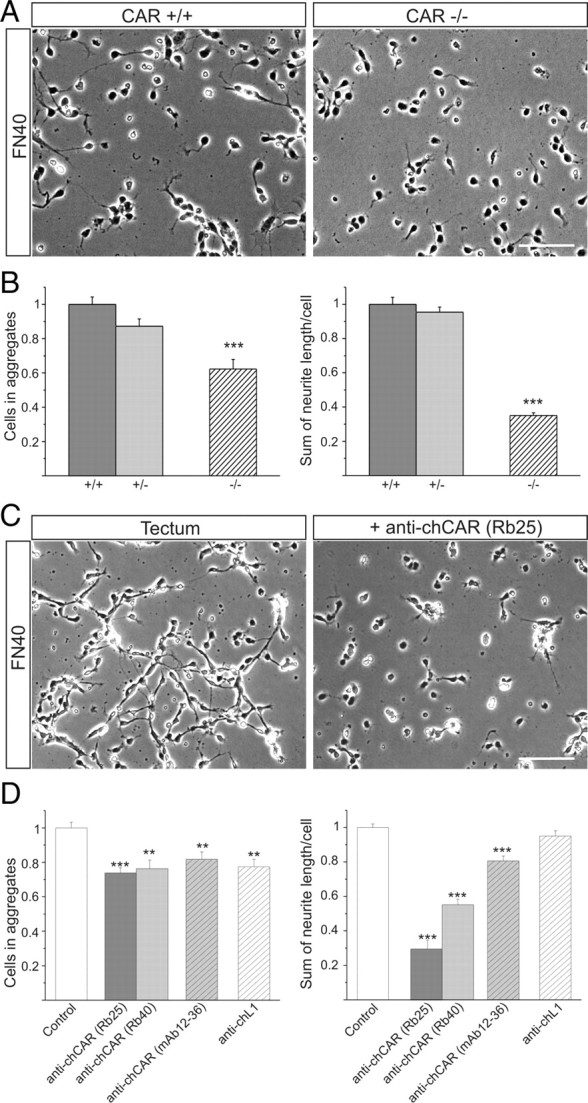
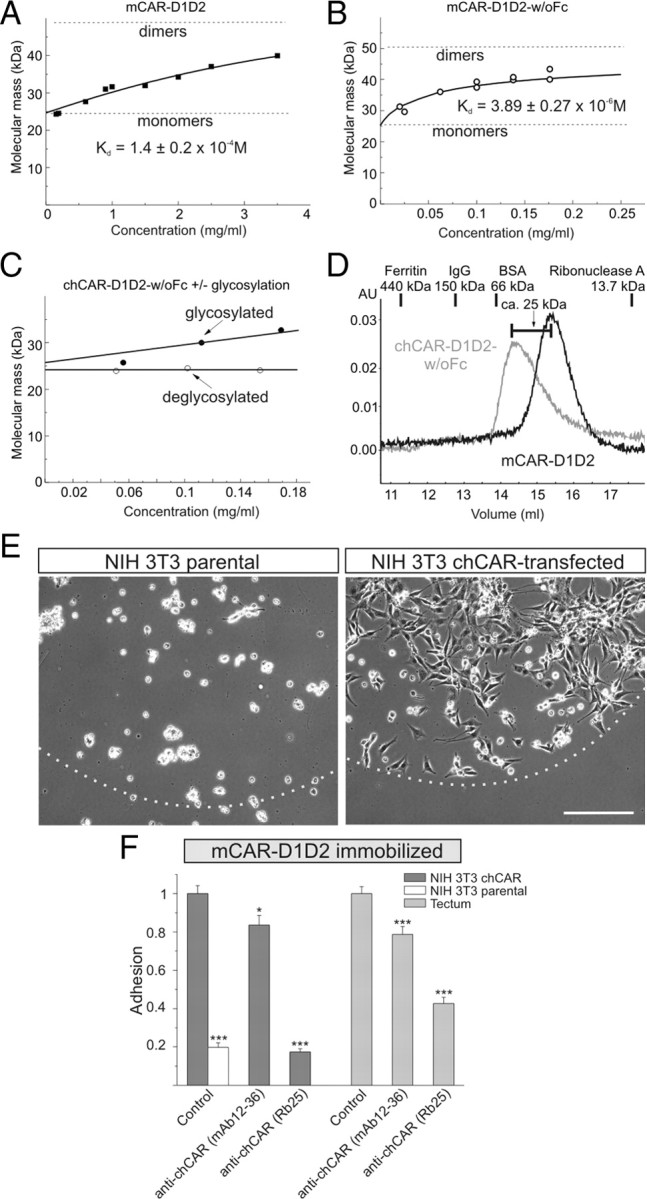
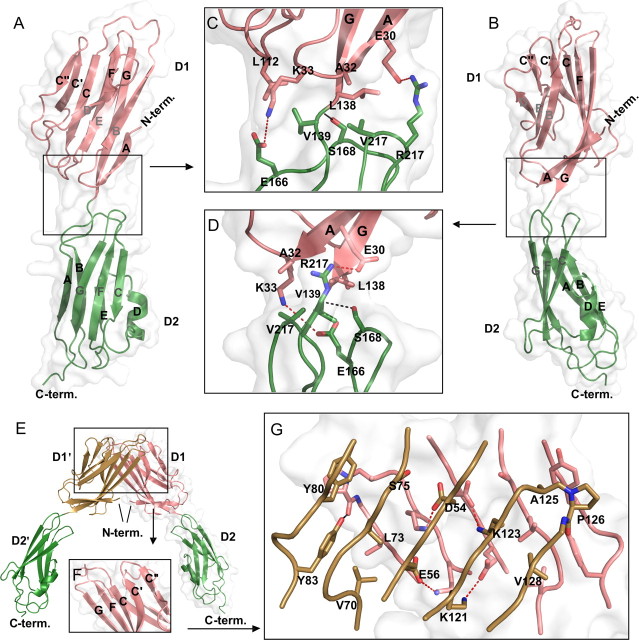
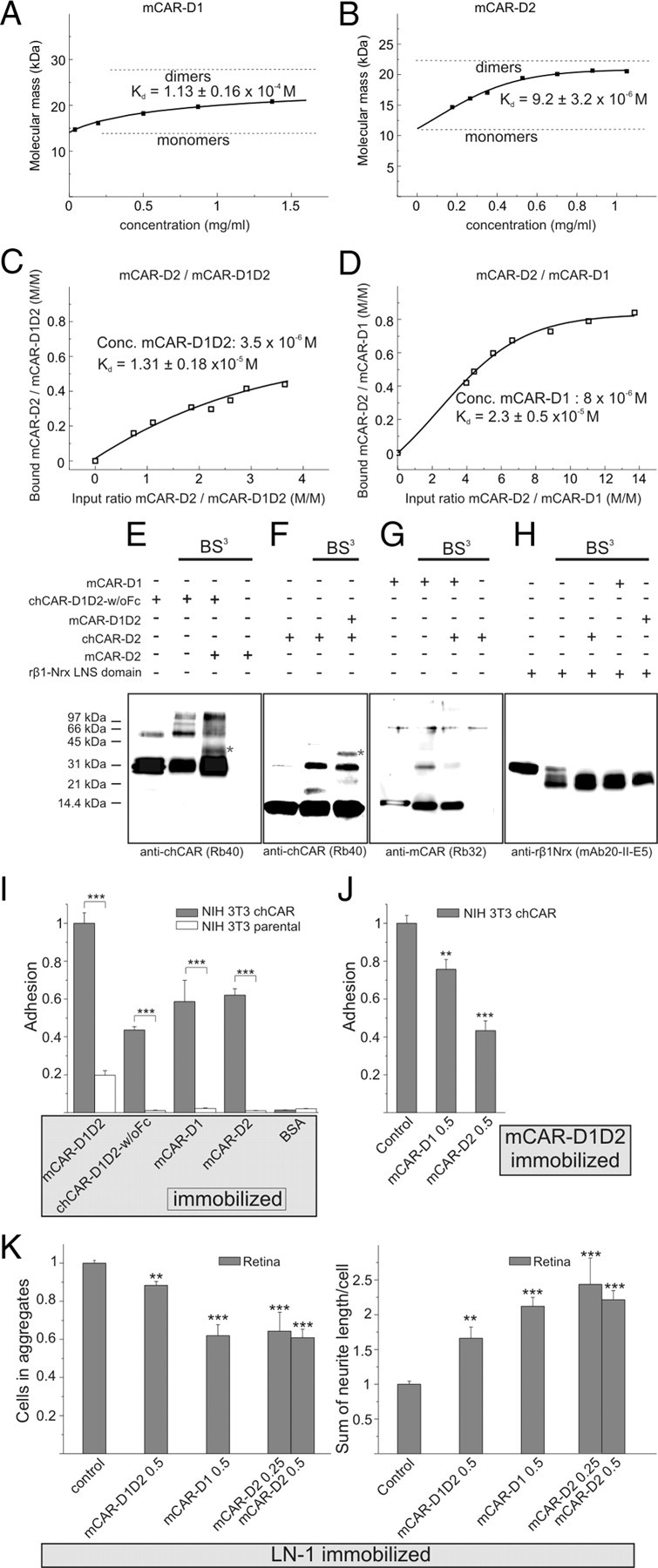
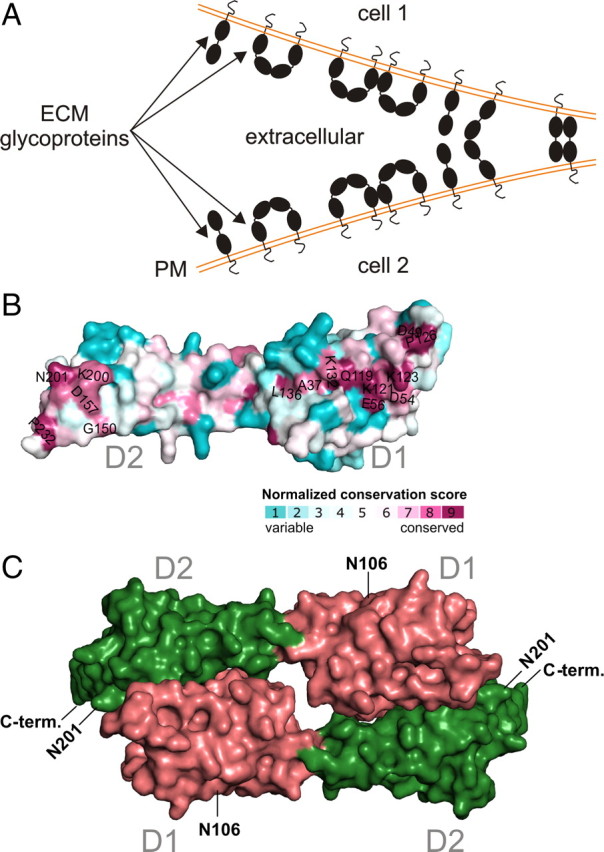
The coxsackievirus-adenovirus receptor (CAR) is a member of the Ig superfamily strongly expressed in the developing nervous system. Our histological investigations during development reveal an initial uniform distribution of CAR on all neural cells with a concentration on membranes that face the margins of the nervous system (e.g., the basal laminae and the ventricular side). At more advanced stages, CAR becomes downregulated and restricted to specific regions including areas rich in axonal and dendritic surfaces. To study the function of CAR on neural cells, we used the fiber knob of the adenovirus, extracellular CAR domains, blocking antibodies to CAR, as well as CAR-deficient neural cells. Blocking antibodies were found to inhibit neurite extension in retina organ and retinal explant cultures, whereas the application of the recombinant fiber knob of the adenovirus subtype Ad2 or extracellular CAR domains promoted neurite extension and adhesion to extracellular matrices. We observed a promiscuous interaction of CAR with extracellular matrix glycoproteins, which was deduced from analytical ultracentrifugation experiments, affinity chromatography, and adhesion assays. The membrane proximal Ig domain of CAR, termed D2, was found to bind to a fibronectin fragment, including the heparin-binding domain 2, which promotes neurite extension of wild type, but not of CAR-deficient neural cells. In contrast to heterophilic interactions, homophilic association of CAR involves both Ig domains, as was revealed by ultracentrifugation, chemical cross-linking, and adhesion studies. The results of these functional and binding studies are correlated to a U-shaped homodimer of the complete extracellular domains of CAR detected by x-ray crystallography.
Figures
References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Asher DR, Cerny AM, Weiler SR, Horner JW, Keeler ML, Neptune MA, Jones SN, Bronson RT, DePinho RA, Finberg RW. Coxsackievirus and adenovirus receptor is essential for cardiomyocyte development. Genesis. 2005;42:77–85. - PubMed
-
- Bailey S. The Ccp4 Suite—Programs for Protein Crystallography. Acta Crystallogr D Biol Crystallogr. 1994;50:760–763. - PubMed
-
- Behlke J, Ristau O, Schonfeld HJ. Nucleotide-dependent complex formation between the Escherichia coli chaperonins GroEL and GroES studied under equilibrium conditions. Biochemistry. 1997;36:5149–5156. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials