

Novel inhibitors of influenza virus fusion: structure-activity relationship and interaction with the viral hemagglutinin
- PMID: 20181685
- PMCID: PMC2863778
- DOI: 10.1128/JVI.02325-09
Novel inhibitors of influenza virus fusion: structure-activity relationship and interaction with the viral hemagglutinin
Abstract
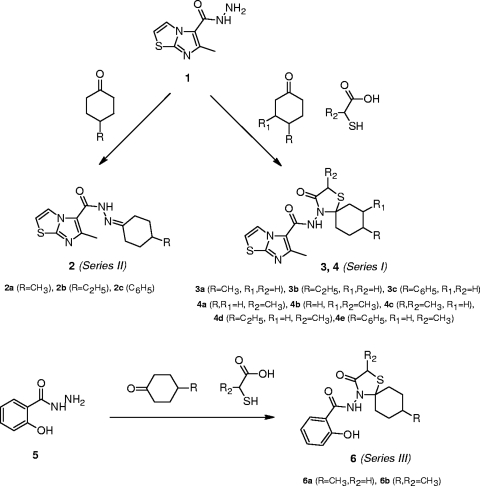
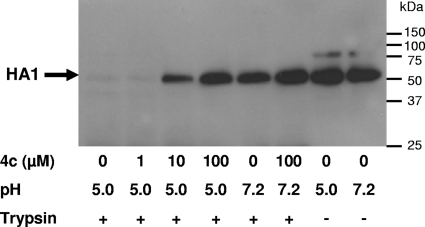
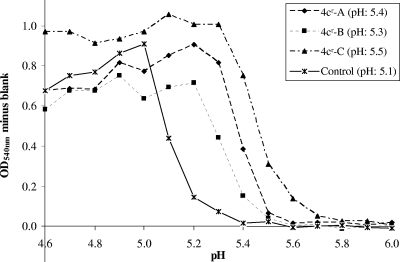
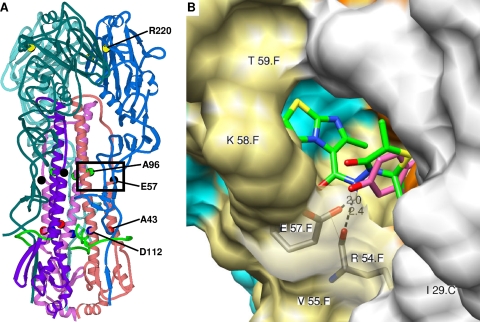
A new class of N-(1-thia-4-azaspiro[4.5]decan-4-yl)carboxamide inhibitors of influenza virus hemagglutinin (HA)-mediated membrane fusion that has a narrow and defined structure-activity relationship was identified. In Madin-Darby canine kidney (MDCK) cells infected with different strains of human influenza virus A/H3N2, the lead compound, 4c, displayed a 50% effective concentration of 3 to 23 muM and an antiviral selectivity index of 10. No activity was observed for A/H1N1, A/H5N1, A/H7N2, and B viruses. The activity of 4c was reduced considerably when added 30 min or later postinfection, indicating that 4c inhibits an early step in virus replication. 4c and its congeners inhibited influenza A/H3N2 virus-induced erythrocyte hemolysis at low pH. 4c-resistant virus mutants, selected in MDCK cells, contained either a single D112N change in the HA2 subunit of the viral HA or a combination of three substitutions, i.e., R220S (in HA1) and E57K (in HA2) and an A-T substitution at position 43 or 96 of HA2. The mutants showed efficiency for receptor binding and replication similar to that of wild-type virus yet displayed an increased pH of erythrocyte hemolysis. In polykaryon assays with cells expressing single-mutant HA proteins, the E57K, A96T, and D112N mutations resulted in 4c resistance, and the HA proteins containing R220S, A96T, and D112N mutations displayed an increased fusion pH. Molecular modeling identified a binding cavity for 4c involving arginine-54 and glutamic acid-57 in the HA2 subunit. Our studies with the new fusion inhibitor 4c confirm the importance of this HA region in the development of influenza virus fusion inhibitors.
Figures


References
-
- Akkurt, M., S. Karaca, E. Sahin, O. Guzel, A. Salman, and E. Ilhan. 2007. 2-Hydroxy-N-(3-oxo-1-thia-4-azaspiro[4.5]dec-4-yl)-2,2-diphenylacetamide. Acta Crystallogr. E. 63:o3379-o3380.
-
- Bodian, D. L., R. B. Yamasaki, R. L. Buswell, J. F. Stearns, J. M. White, and I. D. Kuntz. 1993. Inhibition of the fusion-inducing conformational change of influenza hemagglutinin by benzoquinones and hydroquinones. Biochemistry 32:2967-2978. - PubMed
-
- Bright, R. A., M. J. Medina, X. Xu, G. Perez-Oronoz, T. R. Wallis, X. M. Davis, L. Povinelli, N. J. Cox, and A. I. Klimov. 2005. Incidence of adamantane resistance among influenza A (H3N2) viruses isolated worldwide from 1994 to 2005: a cause for concern. Lancet 366:1175-1181. - PubMed
-
- Bullough, P. A., F. M. Hughson, J. J. Skehel, and D. C. Wiley. 1994. Structure of influenza haemagglutinin at the pH of membrane fusion. Nature 371:37-43. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
