

PB2 residue 271 plays a key role in enhanced polymerase activity of influenza A viruses in mammalian host cells
- PMID: 20181719
- PMCID: PMC2863787
- DOI: 10.1128/JVI.02642-09
PB2 residue 271 plays a key role in enhanced polymerase activity of influenza A viruses in mammalian host cells
Abstract
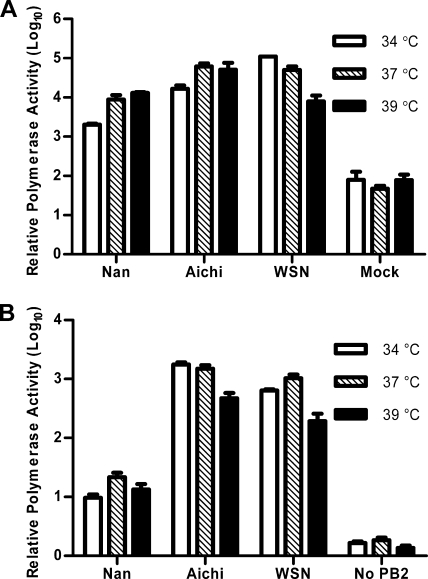
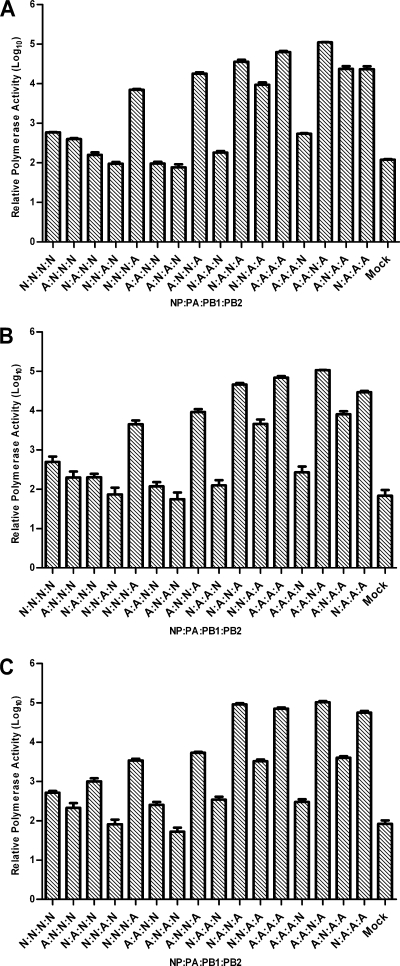
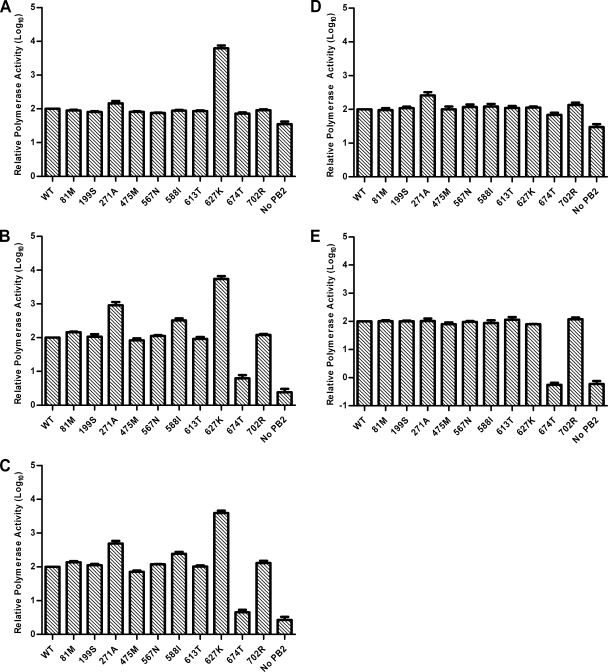
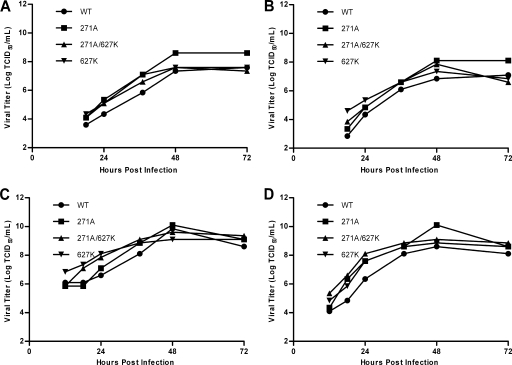
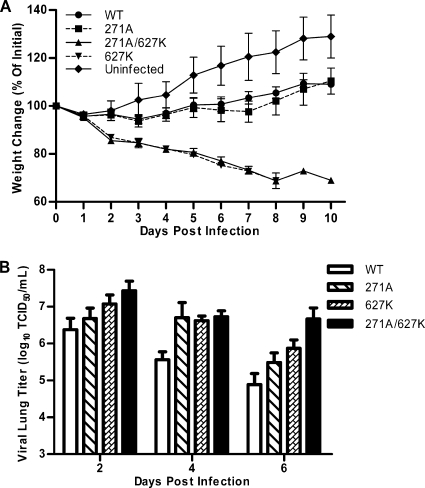
The direct infection of humans with highly pathogenic avian H5N1 influenza viruses has suggested viral mutation as one mechanism for the emergence of novel human influenza A viruses. Although the polymerase complex is known to be a key component in host adaptation, mutations that enhance the polymerase activity of avian viruses in mammalian hosts are not fully characterized. The genomic comparison of influenza A virus isolates has identified highly conserved residues in influenza proteins that are specific to either human or avian viruses, including 10 residues in PB2. We characterized the activity of avian polymerase complexes containing avian-to-human mutations at these conserved PB2 residues and found that, in addition to the E627K mutation, the PB2 mutation T271A enhances polymerase activity in human cells. We confirmed the effects of the T271A mutation using recombinant WSN viruses containing avian NP and polymerase genes with wild-type (WT) or mutant PB2. The 271A virus showed enhanced growth compared to that of the WT in mammalian cells in vitro. The 271A mutant did not increase viral pathogenicity significantly in mice compared to that of the 627K mutant, but it did enhance the lung virus titer. Also, cell infiltration was more evident in lungs of 271A-infected mice than in those of the WT. Interestingly, the avian-derived PB2 of the 2009 pandemic H1N1 influenza virus has 271A. The characterization of the polymerase activity of A/California/04/2009 (H1N1) and corresponding PB2 mutants indicates that the high polymerase activity of the pandemic strain in mammalian cells is, in part, dependent on 271A. Our results clearly indicate the contribution of PB2 amino acid 271 to enhanced polymerase activity and viral growth in mammalian hosts.
Figures
References
-
- Almond, J. W. 1977. A single gene determines the host range of influenza virus. Nature 270:617-618. - PubMed
-
- Butler, D. 2006. Family tragedy spotlights flu mutations. Nature 442:114-115. - PubMed
-
- Claas, E. C., A. D. Osterhaus, R. van Beek, J. C. De Jong, G. F. Rimmelzwaan, D. A. Senne, S. Krauss, K. F. Shortridge, and R. G. Webster. 1998. Human influenza A H5N1 virus related to a highly pathogenic avian influenza virus. Lancet 351:472-477. - PubMed
-
- Dawood, F. S., S. Jain, L. Finelli, M. W. Shaw, S. Lindstrom, R. J. Garten, L. V. Gubareva, X. Xu, C. B. Bridges, and T. M. Uyeki. 2009. Emergence of a novel swine-origin influenza A (H1N1) virus in humans. N Engl. J. Med. 360:2605-2615. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
