

h3/Acidic calponin: an actin-binding protein that controls extracellular signal-regulated kinase 1/2 activity in nonmuscle cells
- PMID: 20181831
- PMCID: PMC2854098
- DOI: 10.1091/mbc.e09-06-0451
h3/Acidic calponin: an actin-binding protein that controls extracellular signal-regulated kinase 1/2 activity in nonmuscle cells
Abstract
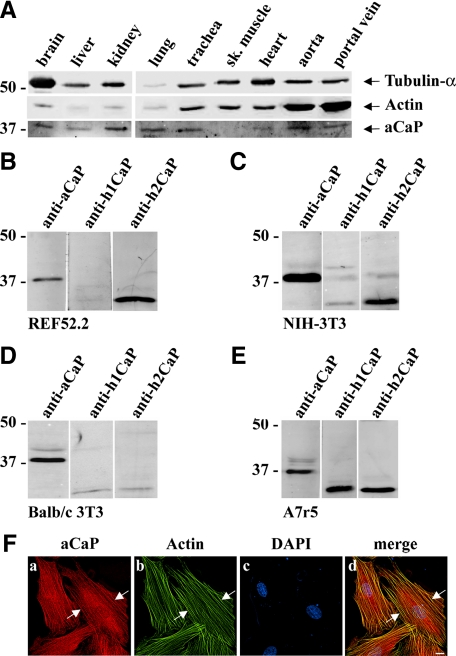
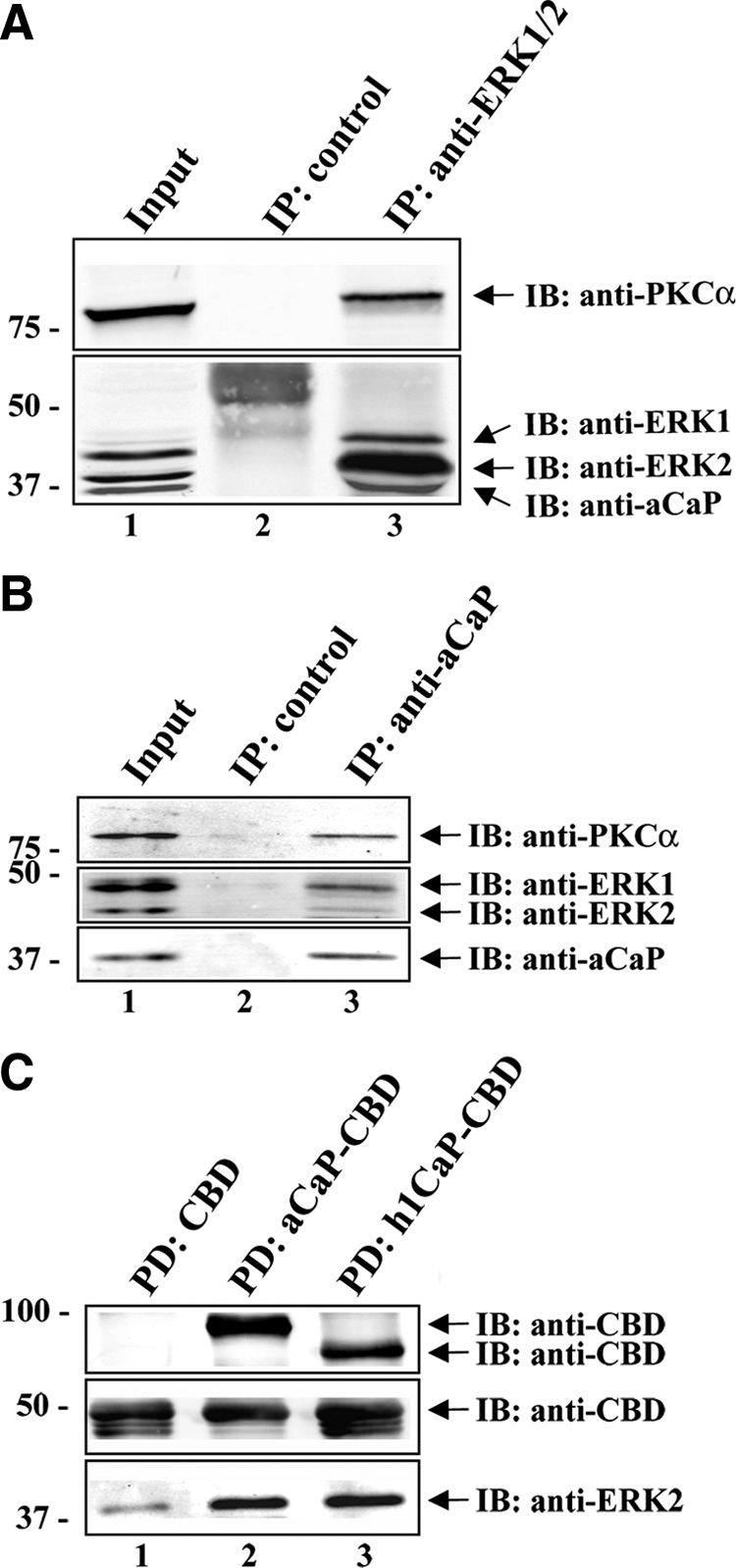
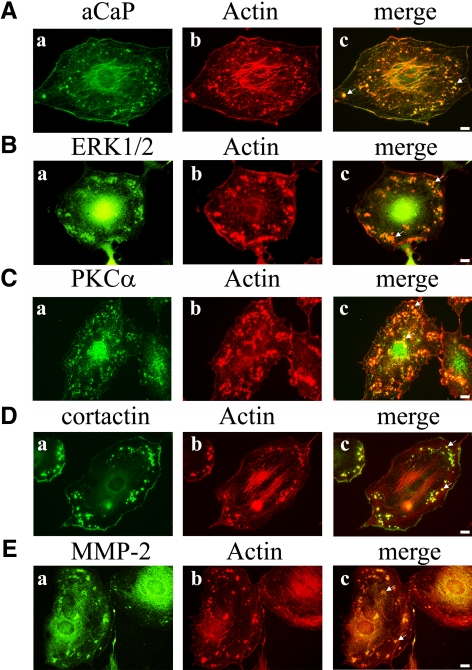
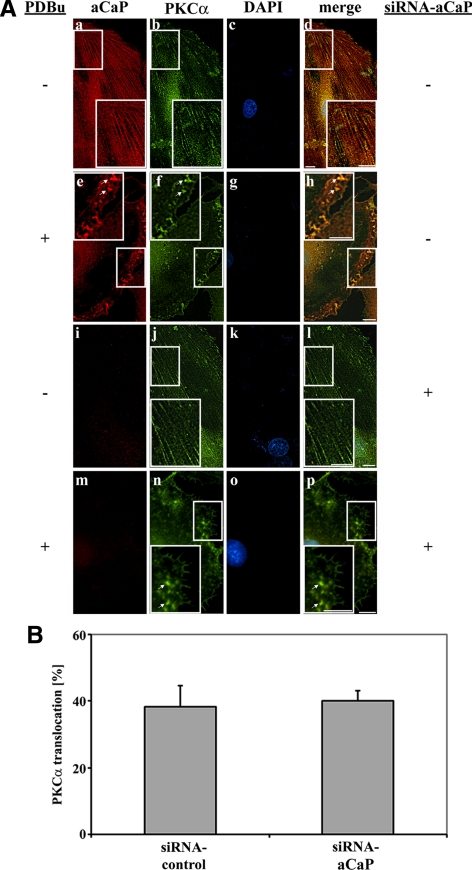
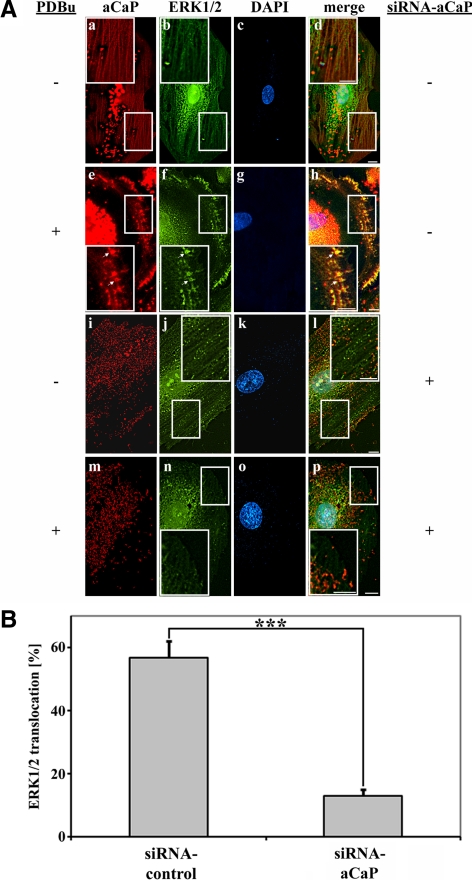
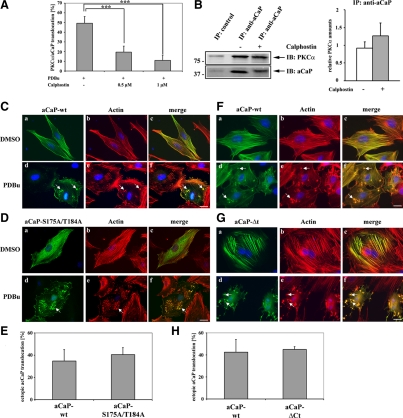
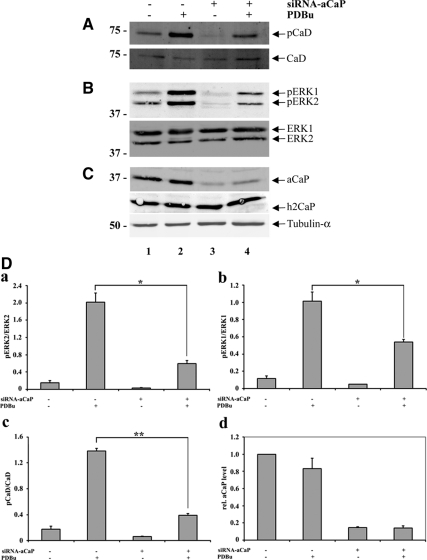
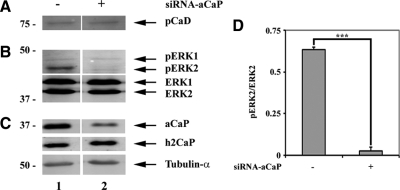
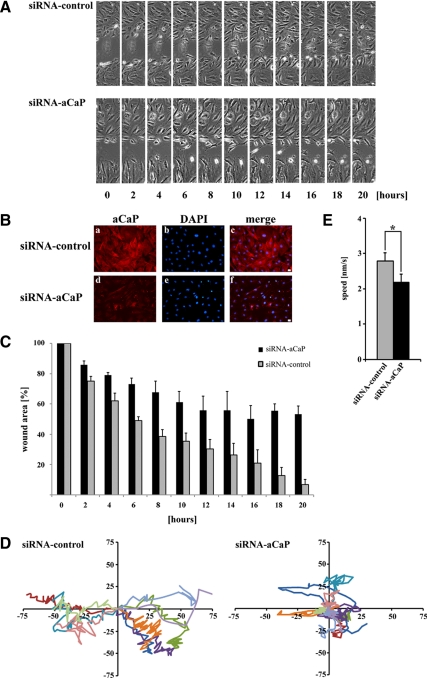
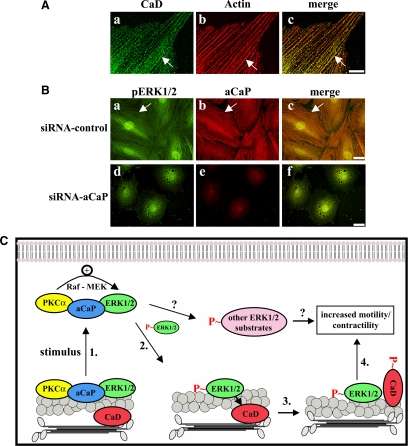
Migration of fibroblasts is important in wound healing. Here, we demonstrate a role and a mechanism for h3/acidic calponin (aCaP, CNN3) in REF52.2 cell motility, a fibroblast line rich in actin filaments. We show that the actin-binding protein h3/acidic calponin associates with stress fibers in the absence of stimulation but is targeted to the cell cortex and podosome-like structures after stimulation with a phorbol ester, phorbol-12,13-dibutyrate (PDBu). By coimmunoprecipitation and colocalization, we show that extracellular signal-regulated kinase (ERK)1/2 and protein kinase C (PKC)alpha constitutively associate with h3/acidic calponin and are cotargeted with h3/acidic calponin in the presence of PDBu. This targeting can be blocked by a PKC inhibitor but does not require phosphorylation of h3/acidic calponin at the PKC sites S175 or T184. Knockdown of h3/acidic calponin results in a loss of PDBu-mediated ERK1/2 targeting, whereas PKCalpha targeting is unaffected. Caldesmon is an actin-binding protein that regulates actomyosin interactions and is a known substrate of ERK1/2. Both ERK1/2 activity and nonmuscle l-caldesmon phosphorylation are blocked by h3/acidic calponin knockdown. Furthermore, h3/acidic calponin knockdown inhibits REF52.2 migration in an in vitro wound healing assay. Our findings are consistent with a model whereby h3/acidic calponin controls fibroblast migration by regulation of ERK1/2-mediated l-caldesmon phosphorylation.
Figures
References
-
- Allen B. G., Walsh M. P. The biochemical basis of the regulation of smooth-muscle contraction. Trends Biochem. Sci. 1994;19:362–368. - PubMed
-
- Applegate D., Feng W., Green R. S., Taubman M. B. Cloning and expression of a novel acidic calponin isoform from rat aortic vascular smooth muscle. J. Biol. Chem. 1994;269:10683–10690. - PubMed
-
- Bruns R. F., Miller F. D., Merriman R. L., Howbert J. J., Heath W. F., Kobayashi E., Takahashi I., Tamaoki T., Nakano H. Inhibition of protein kinase C by calphostin C is light-dependent. Biochem. Biophys. Res. Commun. 1991;176:288–293. - PubMed
-
- Burgstaller G., Kranewitter W. J., Gimona M. The molecular basis for the autoregulation of calponin by isoform-specific C-terminal tail sequences. J. Cell Sci. 2002;115:2021–2029. - PubMed
-
- Cacace A. M., Ueffing M., Philipp A., Han E. K., Kolch W., Weinstein I. B. PKC epsilon functions as an oncogene by enhancing activation of the Raf kinase. Oncogene. 1996;13:2517–2526. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
