

Matrix metalloproteinase 12-deficiency augments extracellular matrix degrading metalloproteinases and attenuates IL-13-dependent fibrosis
- PMID: 20181883
- PMCID: PMC3175622
- DOI: 10.4049/jimmunol.0903008
Matrix metalloproteinase 12-deficiency augments extracellular matrix degrading metalloproteinases and attenuates IL-13-dependent fibrosis
Abstract
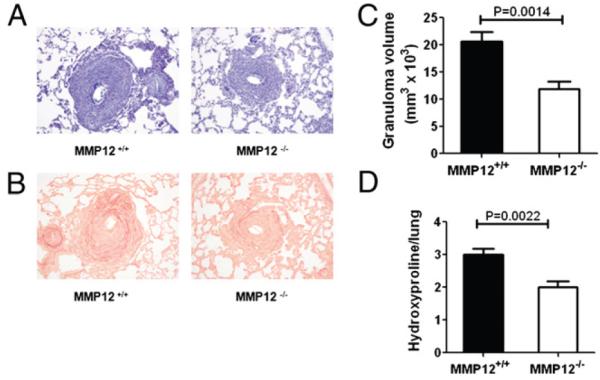
Infection with the parasitic helminth Schistosoma mansoni causes significant liver fibrosis and extracellular matrix (ECM) remodeling. Matrix metalloproteinases (MMP) are important regulators of the ECM by regulating cellular inflammation, extracellular matrix deposition, and tissue reorganization. MMP12 is a macrophage-secreted elastase that is highly induced in the liver and lung in response to S. mansoni eggs, confirmed by both DNA microarray and real-time PCR analysis. However, the function of MMP12 in chronic helminth-induced inflammation and fibrosis is unclear. In this study, we reveal that MMP12 acts as a potent inducer of inflammation and fibrosis after infection with the helminth parasite S. mansoni. Surprisingly, the reduction in liver and lung fibrosis in MMP12-deficient mice was not associated with significant changes in cytokine, chemokine, TGF-beta1, or tissue inhibitors of matrix metalloproteinase expression. Instead, we observed marked increases in MMP2 and MMP13 expression, suggesting that Mmp12 was promoting fibrosis by limiting the expression of specific ECM-degrading MMPs. Interestingly, like MMP12, MMP13 expression was highly dependent on IL-13 and type II-IL-4 receptor signaling. However, in contrast to MMP12, expression of MMP13 was significantly suppressed by the endogenous IL-13 decoy receptor, IL-13Ralpha2. In the absence of MMP12, expression of IL-13Ralpha2 was significantly reduced, providing a possible explanation for the increased IL-13-driven MMP13 activity and reduced fibrosis. As such, these data suggest important counter-regulatory roles between MMP12 and ECM-degrading enzymes like MMP2, MMP9, and MMP13 in Th2 cytokine-driven fibrosis.
Figures






References
-
- Tallant C, Marrero A, Gomis-Ruth FX. Matrix metalloproteinases: Fold and function of their catalytic domains. Biochim Biophys Acta - Molecular. Cell Res. 2010;1803:20–28. - PubMed
-
- Vaillant B, Chiaramonte MG, Cheever AW, Soloway PD, Wynn TA. Regulation of hepatic fibrosis and extracellular matrix genes by the th response: new insight into the role of tissue inhibitors of matrix metalloproteinases. J. Immunol. 2001;167:7017–7026. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
