

Adenosine A1 receptors (A1Rs) play a critical role in osteoclast formation and function
- PMID: 20181934
- PMCID: PMC2887264
- DOI: 10.1096/fj.09-147447
Adenosine A1 receptors (A1Rs) play a critical role in osteoclast formation and function
Abstract
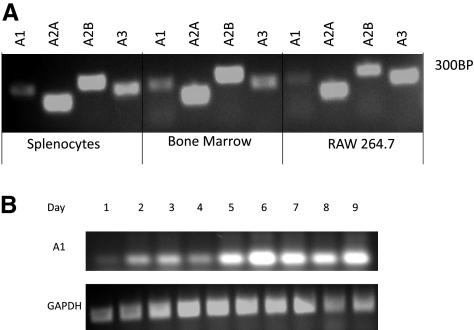
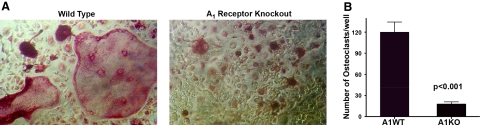
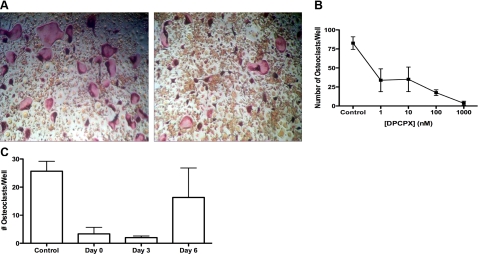
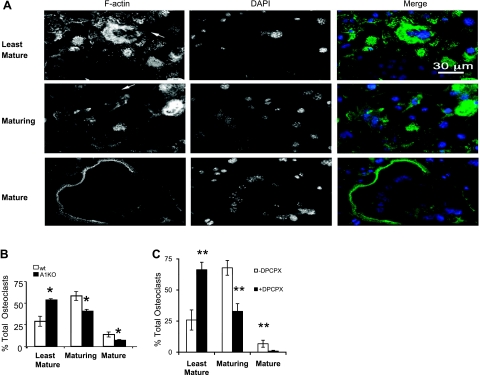
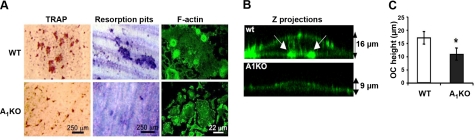
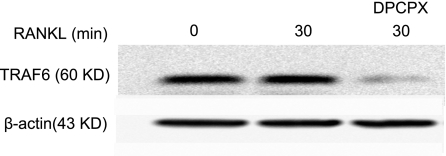
Adenosine regulates a wide variety of physiological processes via interaction with one or more G-protein-coupled receptors (A(1)R, A(2A)R, A(2B)R, and A(3)R). Because A(1)R occupancy promotes fusion of human monocytes to form giant cells in vitro, we determined whether A(1)R occupancy similarly promotes osteoclast function and formation. Bone marrow cells (BMCs) were harvested from C57Bl/6 female mice or A(1)R-knockout mice and their wild-type (WT) littermates and differentiated into osteoclasts in the presence of colony stimulating factor-1 and receptor activator of NF-kappaB ligand in the presence or absence of the A(1)R antagonist 1,3-dipropyl-8-cyclopentyl xanthine (DPCPX). Osteoclast morphology was analyzed in tartrate-resistant acid phosphatase or F-actin-stained samples, and bone resorption was evaluated by toluidine blue staining of dentin. BMCs from A(1)R-knockout mice form fewer osteoclasts than BMCs from WT mice, and the A(1)R antagonist DPCPX inhibits osteoclast formation (IC(50)=1 nM), with altered morphology and reduced ability to resorb bone. A(1)R blockade increased ubiquitination and degradation of TRAF6 in RAW264.7 cells induced to differentiate into osteoclasts. These studies suggest a critical role for adenosine in bone homeostasis via interaction with adenosine A(1)R and further suggest that A(1)R may be a novel pharmacologic target to prevent the bone loss associated with inflammatory diseases and menopause.
Figures
Similar articles
-
Rolofylline, an adenosine A1 receptor antagonist, inhibits osteoclast differentiation as an inverse agonist.Br J Pharmacol. 2013 Nov;170(6):1167-76. doi: 10.1111/bph.12342. Br J Pharmacol. 2013. PMID: 23962057 Free PMC article.
-
Shikimic Acid Inhibits Osteoclastogenesis in Vivo and in Vitro by Blocking RANK/TRAF6 Association and Suppressing NF-κB and MAPK Signaling Pathways.Cell Physiol Biochem. 2018;51(6):2858-2871. doi: 10.1159/000496039. Epub 2018 Dec 14. Cell Physiol Biochem. 2018. PMID: 30562759
-
Tumor necrosis factor-alpha induces differentiation of and bone resorption by osteoclasts.J Biol Chem. 2000 Feb 18;275(7):4858-64. doi: 10.1074/jbc.275.7.4858. J Biol Chem. 2000. PMID: 10671521
-
Differentiation and function of osteoclasts.Keio J Med. 2003 Mar;52(1):1-7. doi: 10.2302/kjm.52.1. Keio J Med. 2003. PMID: 12713016 Review.
-
The roles of adenosine and adenosine receptors in bone remodeling.Front Biosci (Elite Ed). 2011 Jun 1;3(3):888-95. doi: 10.2741/e297. Front Biosci (Elite Ed). 2011. PMID: 21622100 Review.
Cited by
-
Lack of effect of adenosine on the function of rodent osteoblasts and osteoclasts in vitro.Purinergic Signal. 2016 Jun;12(2):247-58. doi: 10.1007/s11302-016-9499-2. Epub 2016 Feb 10. Purinergic Signal. 2016. PMID: 26861849 Free PMC article.
-
Caffeine displays dual effect on RANKL-induced osteoclast differentiation and bone resorption activity via adenosine receptors in RAW 264.7 cells.J Orofac Orthop. 2025 Aug 19. doi: 10.1007/s00056-025-00611-z. Online ahead of print. J Orofac Orthop. 2025. PMID: 40828391 English.
-
Regulation of bone and cartilage by adenosine signaling.Purinergic Signal. 2016 Dec;12(4):583-593. doi: 10.1007/s11302-016-9527-2. Epub 2016 Jul 29. Purinergic Signal. 2016. PMID: 27473363 Free PMC article. Review.
-
Bone Marrow Adipose Tissue and Skeletal Health.Curr Osteoporos Rep. 2018 Aug;16(4):434-442. doi: 10.1007/s11914-018-0451-y. Curr Osteoporos Rep. 2018. PMID: 29855795 Free PMC article. Review.
-
Signaling of the Purinergic System in the Joint.Front Pharmacol. 2020 Jan 24;10:1591. doi: 10.3389/fphar.2019.01591. eCollection 2019. Front Pharmacol. 2020. PMID: 32038258 Free PMC article. Review.
References
-
- Finkelstein J S, Dawson-Hugues B. Osteoporosis. Goldman L, Bennett J C, editors. Philadelphia: Saunders; Cecil Textbook of Medicine. (21st ed) 2000:1366–1373.
-
- Rodan G A, Martin T J. Therapeutic approaches to bone diseases. Science. 2000;289:1508–1514. - PubMed
-
- Suda T, Takahashi N, Udagawa N, Jimi E, Gillespie M T, Martin T J. Modulation of osteoclast differentiation and function by the new members of the tumor necrosis factor receptor and ligand families. Endocr Rev. 1999;20:345–357. - PubMed
-
- Karsenty G, Wagner E F. Reaching a genetic and molecular understanding of skeletal development. Dev Cell. 2002;2:389–406. - PubMed
-
- Manolagas S C. Birth and death of bone cells: basic regulatory mechanisms and implications for the pathogenesis and treatment of osteoporosis. Endocr Rev. 2000;21:115–137. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 AR054897/AR/NIAMS NIH HHS/United States
- K01 AR054486-02/AR/NIAMS NIH HHS/United States
- R01 CA032551/CA/NCI NIH HHS/United States
- UL1 RR029893/RR/NCRR NIH HHS/United States
- AA13336/AA/NIAAA NIH HHS/United States
- R37 CA026504/CA/NCI NIH HHS/United States
- UL1RR029893/RR/NCRR NIH HHS/United States
- R01 AR041911/AR/NIAMS NIH HHS/United States
- R01 CA25604/CA/NCI NIH HHS/United States
- AR41911/AR/NIAMS NIH HHS/United States
- P30 AR046121/AR/NIAMS NIH HHS/United States
- K01 AR054486/AR/NIAMS NIH HHS/United States
- AR54897/AR/NIAMS NIH HHS/United States
- P30 CA013330/CA/NCI NIH HHS/United States
- R01 AA013336/AA/NIAAA NIH HHS/United States
- P30 CA13330/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
