

Polarity in stem cell division: asymmetric stem cell division in tissue homeostasis
- PMID: 20182603
- PMCID: PMC2827902
- DOI: 10.1101/cshperspect.a001313
Polarity in stem cell division: asymmetric stem cell division in tissue homeostasis
Abstract
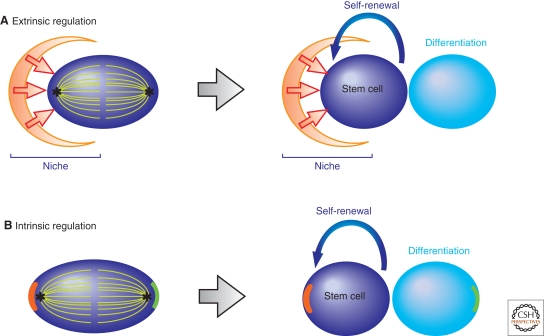
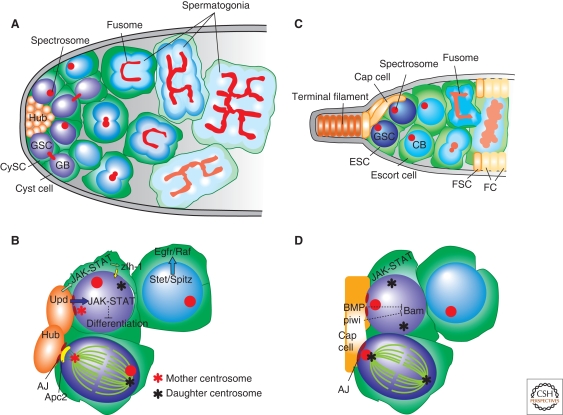
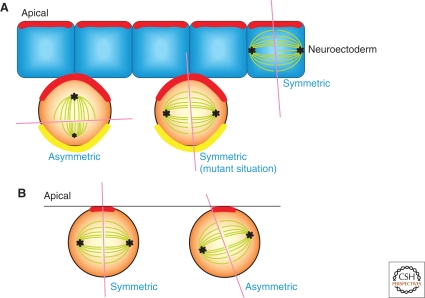
Many adult stem cells divide asymmetrically to balance self-renewal and differentiation, thereby maintaining tissue homeostasis. Asymmetric stem cell divisions depend on asymmetric cell architecture (i.e., cell polarity) within the cell and/or the cellular environment. In particular, as residents of the tissues they sustain, stem cells are inevitably placed in the context of the tissue architecture. Indeed, many stem cells are polarized within their microenvironment, or the stem cell niche, and their asymmetric division relies on their relationship with the microenvironment. Here, we review asymmetric stem cell divisions in the context of the stem cell niche with a focus on Drosophila germ line stem cells, where the nature of niche-dependent asymmetric stem cell division is well characterized.
Figures
Similar articles
-
Cellular and molecular mechanisms of asymmetric stem cell division in tissue homeostasis.Genes Cells. 2024 Dec;29(12):1099-1110. doi: 10.1111/gtc.13172. Epub 2024 Oct 8. Genes Cells. 2024. PMID: 39379096 Free PMC article. Review.
-
Asymmetric stem cell division and function of the niche in the Drosophila male germ line.Int J Hematol. 2005 Dec;82(5):377-80. doi: 10.1532/IJH97.05097. Int J Hematol. 2005. PMID: 16533738 Review.
-
Asymmetric inheritance of mother versus daughter centrosome in stem cell division.Science. 2007 Jan 26;315(5811):518-21. doi: 10.1126/science.1134910. Science. 2007. PMID: 17255513 Free PMC article.
-
The ins(ide) and outs(ide) of asymmetric stem cell division.Curr Opin Cell Biol. 2016 Dec;43:1-6. doi: 10.1016/j.ceb.2016.06.001. Epub 2016 Jun 16. Curr Opin Cell Biol. 2016. PMID: 27318429 Free PMC article. Review.
-
The polarity protein Baz forms a platform for the centrosome orientation during asymmetric stem cell division in the Drosophila male germline.Elife. 2015 Mar 20;4:e04960. doi: 10.7554/eLife.04960. Elife. 2015. PMID: 25793442 Free PMC article.
Cited by
-
Age-related changes in the hematopoietic stem cell pool revealed via quantifying the balance of symmetric and asymmetric divisions.PLoS One. 2024 Jan 29;19(1):e0292575. doi: 10.1371/journal.pone.0292575. eCollection 2024. PLoS One. 2024. PMID: 38285676 Free PMC article.
-
Aging alters the epigenetic asymmetry of HSC division.PLoS Biol. 2018 Sep 20;16(9):e2003389. doi: 10.1371/journal.pbio.2003389. eCollection 2018 Sep. PLoS Biol. 2018. PMID: 30235201 Free PMC article.
-
Eng2, a new player involved in feedback loop regulation of Cdc42 activity in fission yeast.Sci Rep. 2021 Sep 9;11(1):17872. doi: 10.1038/s41598-021-97311-6. Sci Rep. 2021. PMID: 34504165 Free PMC article.
-
Spatial control of translation repression and polarized growth by conserved NDR kinase Orb6 and RNA-binding protein Sts5.Elife. 2016 Jul 30;5:e14216. doi: 10.7554/eLife.14216. Elife. 2016. PMID: 27474797 Free PMC article.
-
A comprehensive review of computational cell cycle models in guiding cancer treatment strategies.NPJ Syst Biol Appl. 2024 Jul 5;10(1):71. doi: 10.1038/s41540-024-00397-7. NPJ Syst Biol Appl. 2024. PMID: 38969664 Free PMC article. Review.
References
-
- Akong K, McCartney B, Peifer M 2002b. Drosophila APC2 and APC1 have overlapping roles in the larval brain despite their distinct intracellular localizations. Dev Biol 250:71. - PubMed
-
- Akong K, Grevengoed E, Price M, McCartney B, Hayden M, DeNofrio J, Peifer M 2002a. Drosophila APC2 and APC1 play overlapping roles in wingless signaling in the embryo and imaginal discs. Dev Biol 250:91. - PubMed
-
- Asaoka M, Lin H 2004. Germline stem cells in the Drosophila ovary descend from pole cells in the anterior region of the embryonic gonad. Development 131:5079–5089 - PubMed
-
- Boyle M, Wong C, Rocha M, Jones DL 2007. Decline in self-renewal factors contributes to aging of the stem cell niche in the Drosophila testis. Cell Stem Cell 1:470–478 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases