

Control of leaf and vein development by auxin
- PMID: 20182604
- PMCID: PMC2827905
- DOI: 10.1101/cshperspect.a001511
Control of leaf and vein development by auxin
Abstract
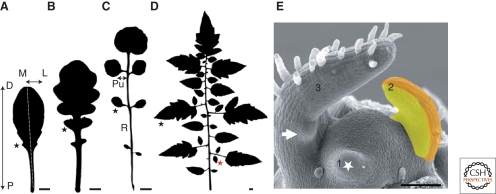
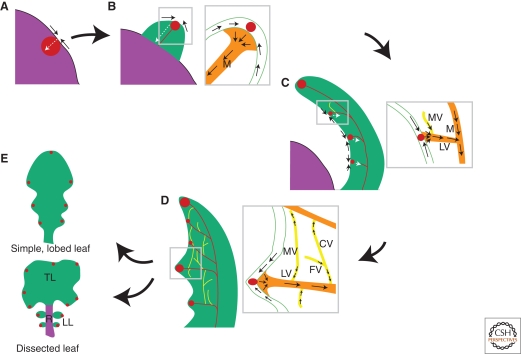
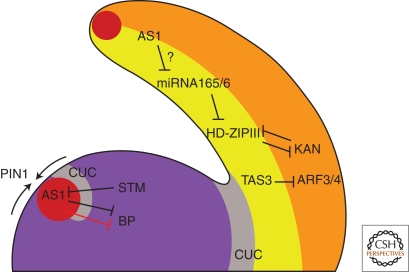
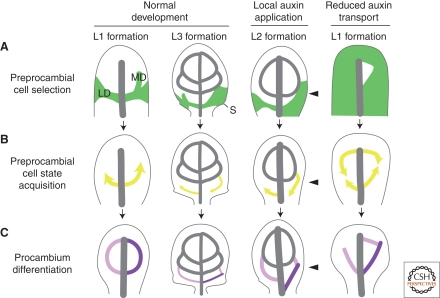
Leaves are the main photosynthetic organs of vascular plants and show considerable diversity in their geometries, ranging from simple spoon-like forms to complex shapes with individual leaflets, as in compound leaves. Leaf vascular tissues, which act as conduits of both nutrients and signaling information, are organized in networks of different architectures that usually mirror the surrounding leaf shape. Understanding the processes that endow leaves and vein networks with ordered and closely aligned shapes has captured the attention of biologists and mathematicians since antiquity. Recent work has suggested that the growth regulator auxin has a key role in both initiation and elaboration of final morphology of both leaves and vascular networks. A key feature of auxin action is the existence of feedback loops through which auxin regulates its own transport. These feedbacks may facilitate the iterative generation of basic modules that underlies morphogenesis of both leaves and vasculature.
Figures
References
-
- Aida M, Vernoux T, Furutani M, Traas J, Tasaka M 2002. Roles of PIN-FORMED1 and MONOPTEROS in pattern formation of the apical region of the Arabidopsis embryo. Development 129:3965–3974 - PubMed
-
- Aloni R, Schwalm K, Langhans M, Ullrich CI 2003. Gradual shifts in sites of free-auxin production during leaf-primordium development and their role in vascular differentiation and leaf morphogenesis in Arabidopsis. Planta 216:841–853 - PubMed
-
- Alonso-Peral MM, Candela H, del Pozo JC, Martinez-Laborda A, Ponce MR, Micol JL 2006. The HVE/CAND1 gene is required for the early patterning of leaf venation in Arabidopsis. Deveopment 133:3755–3766 - PubMed
-
- Avasarala S, Wang J, Caruso JL 1996. Production of phenocopies of the lanceolate mutant in tomato using polar auxin transport inhibitors. J Exp Bot 47:709–712
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources