

Review
doi: 10.1101/cshperspect.a001669.
Regulation of organ growth by morphogen gradients
Affiliations
- PMID: 20182606
- PMCID: PMC2827898
- DOI: 10.1101/cshperspect.a001669
Item in Clipboard
Review
Regulation of organ growth by morphogen gradients
Cold Spring Harb Perspect Biol.
2010 Jan.
Abstract
Morphogen gradients play a fundamental role in organ patterning and organ growth. Unlike their role in patterning, their function in regulating the growth and the size of organs is poorly understood. How and why do morphogen gradients exert their mitogenic effects to generate uniform proliferation in developing organs, and by what means can morphogens impinge on the final size of organs? The decapentaplegic (Dpp) gradient in the Drosophila wing imaginal disc has emerged as a suitable and established system to study organ growth. Here, we review models and recent findings that attempt to address how the Dpp morphogen contributes to uniform proliferation of cells, and how it may regulate the final size of wing discs.
Figures
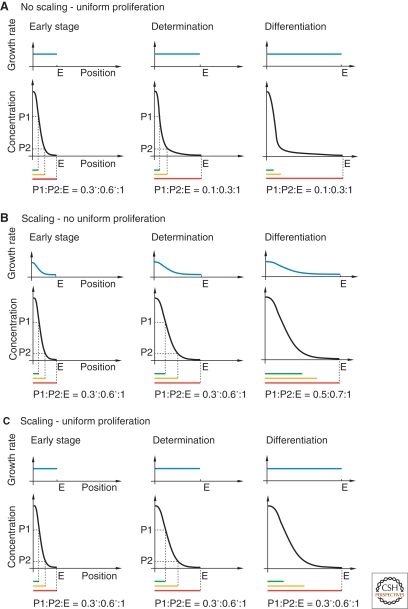
Uniform growth together with scaled morphogen gradients allows determination of cell fates at different time points without losing the right proportions. (A) The case in which growth is uniform, but the morphogen gradient is not scaled. This leads to a change in the proportions of target gene expression domains. Thus, in this situation, the final proportions strictly depend on the precise time-point of cell fate determination. (B) A morphogen gradient scaled to organ size, but driving nonuniform growth. The expression domains of target genes are in proportion only until cell fate determination. Thus, also here, the time point of cell fate determination cannot be shifted without a change in the final proportions. (C) A morphogen gradient scaled to organ size together with uniform growth. In this situation, the expression domains of target genes keep the right proportions independent of the time-point of cell fate determination. Note that the values of the proportions are not based on experimental data. (P1 and P2) Threshold levels for target gene 1 and 2, (E) endpoint of the organ.
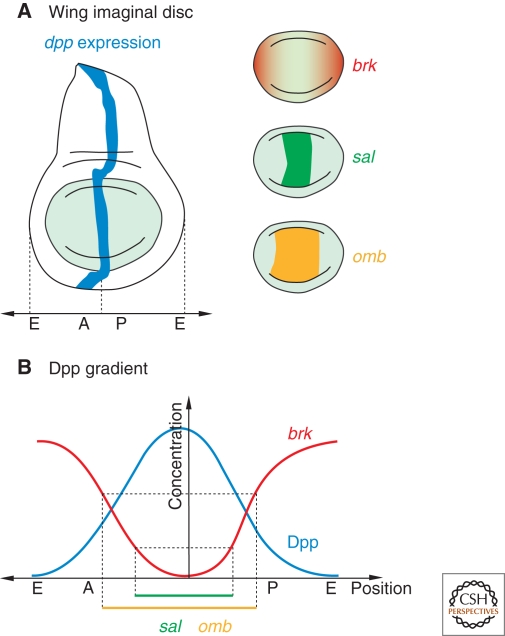
The Dpp gradient in the developing wing imaginal disc of Drosophila serves as a model to study organ patterning and growth. (A) Expression domains of dpp, brk, sal, and omb. (B) Dpp is secreted from its site of production in the center of the disc and spreads into the A and P compartment, establishing a gradient with highest levels in the center and lowest in the periphery. brk transcription is gradually repressed by Dpp signaling activity, leading to a brk gradient inverse to the Dpp gradient. Brk levels are important to set the expression boundaries of sal and omb. omb is repressed by high levels of Brk, and sal by lower levels. Consequently, omb is expressed in a broader domain than sal. (A) Anterior compartment, (P) posterior compartment.
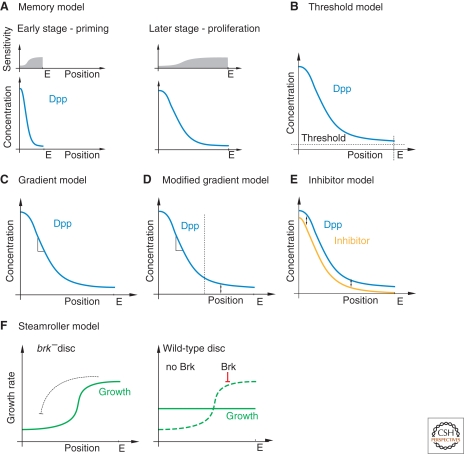
Models addressing how the Dpp gradient can account for uniform growth within the wing disc. (A) Cells are primed for different sensitivity to the mitogenic activity of Dpp, which leads to uniform proliferation during the growth period. (B) Dpp drives proliferation at constant levels above the threshold value, which is reached within the entire wing disc. (C) In the gradient model, the slope of the Dpp gradient drives proliferation. (D) A modified version of the gradient model, in which the disc is subdivided into two regions, the medial and the lateral regions. In the medial region, where the Dpp gradient is sufficiently steep, cells proliferate in response to the gradient, and in the lateral region, where the gradient becomes very shallow, cells proliferate in response to absolute Dpp levels. (E) A growth inhibitor in parallel to Dpp evens out high differences of Dpp signaling along the AP axis. (F) In this model, lateral cells have a growth advantage compared with medial cells, and the Dpp/Brk system is needed to curb lateral proliferation. If absent, lateral overproliferation by an unknown mechanism inhibits growth in the medial area.
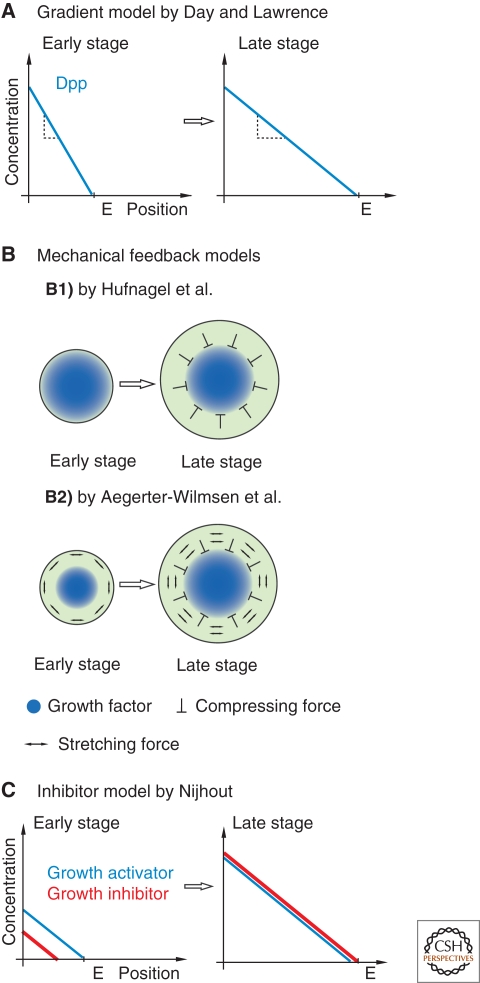
Models for final size control of the wing disc by the Dpp gradient. (A) This model is based on the assumption that Dpp levels in the center and the periphery are fixed, resulting in a decline of the slope of the Dpp gradient when the disc grows. Growth stops when the slope is sufficiently shallow. (B1) This model assumes that the Dpp gradient does not adapt to the disc size. Once the disc has grown, lateral cells do not receive sufficient Dpp and stop proliferating. A mechanical feedback mechanism exerts a compression force to medial cells, leading to a growth stop also in this region. (B2) Growth of the wing disc is governed by a combination of morphogens, which are concentrated at the center of the disc, and stretching and compression forces. High growth factor levels in the center of the disc lead to local growth. This causes stretching, and thus growth in the circumferential regions of the disc, with some stretching still remaining. Stretching forces in the periphery in turn lead to compression of medial cells, which influences growth negatively. As the disc grows, the stretched region becomes wider and compression in the center of the disc becomes stronger. Growth stops once the influence of compression forces becomes bigger than the growth promoting influence by Dpp. (C) In parallel to a growth activator, a growth inhibitor is expressed. Initially, the growth activator is expressed at higher levels. Over time, this ratio changes, and growth stops when the inhibitor reaches a concentration higher than the activator.
Similar articles
-
Dynamics of Dpp signaling and proliferation control.Science. 2011 Mar 4;331(6021):1154-9. doi: 10.1126/science.1200037. Science. 2011. PMID: 21385708
-
Is Drosophila Dpp/BMP morphogen spreading required for wing patterning and growth?Bioessays. 2023 Sep;45(9):e2200218. doi: 10.1002/bies.202200218. Epub 2023 Jul 14. Bioessays. 2023. PMID: 37452394 Review.
-
Scaling morphogen gradients during tissue growth by a cell division rule.Development. 2014 May;141(10):2150-6. doi: 10.1242/dev.107011. Development. 2014. PMID: 24803660
-
BMP morphogen gradients in flies.Cytokine Growth Factor Rev. 2016 Feb;27:119-27. doi: 10.1016/j.cytogfr.2015.11.003. Epub 2015 Nov 24. Cytokine Growth Factor Rev. 2016. PMID: 26684043
-
The wing and the eye: a parsimonious theory for scaling and growth control?Wiley Interdiscip Rev Dev Biol. 2015 Nov-Dec;4(6):591-608. doi: 10.1002/wdev.195. Epub 2015 Jun 24. Wiley Interdiscip Rev Dev Biol. 2015. PMID: 26108346 Review.
Cited by
-
Dpp spreading is required for medial but not for lateral wing disc growth.Nature. 2015 Nov 19;527(7578):317-22. doi: 10.1038/nature15712. Epub 2015 Nov 9. Nature. 2015. PMID: 26550827
-
Decapentaplegic (dpp) regulates the growth of a morphological novelty, beetle horns.Dev Genes Evol. 2011 May;221(1):17-27. doi: 10.1007/s00427-011-0355-7. Epub 2011 Mar 12. Dev Genes Evol. 2011. PMID: 21399983
-
Mechanosensitive mechanisms in transcriptional regulation.J Cell Sci. 2012 Jul 1;125(Pt 13):3061-73. doi: 10.1242/jcs.093005. Epub 2012 Jul 13. J Cell Sci. 2012. PMID: 22797927 Free PMC article. Review.
-
The Hippo Signaling Pathway in Development and Disease.Dev Cell. 2019 Aug 5;50(3):264-282. doi: 10.1016/j.devcel.2019.06.003. Dev Cell. 2019. PMID: 31386861 Free PMC article. Review.
-
Modern perspectives on near-equilibrium analysis of Turing systems.Philos Trans A Math Phys Eng Sci. 2021 Dec 27;379(2213):20200268. doi: 10.1098/rsta.2020.0268. Epub 2021 Nov 8. Philos Trans A Math Phys Eng Sci. 2021. PMID: 34743603 Free PMC article. Review.
References
-
- Adachi-Yamada T, O'Connor MB 2002. Morphogenetic apoptosis: A mechanism for correcting discontinuities in morphogen gradients. Dev Biol 251:74–90 - PubMed
-
- Adachi-Yamada T, Fujimura-Kamada K, Nishida Y, Matsumoto K 1999. Distortion of proximodistal information causes JNK-dependent apoptosis in Drosophila wing. Nature 400:166–169 - PubMed
-
- Aegerter-Wilmsen T, Aegerter CM, Hafen E, Basler K 2007. Model for the regulation of size in the wing imaginal disc of Drosophila. Mech Dev 124:318–326 - PubMed
-
- Affolter M, Basler K 2007. The Decapentaplegic morphogen gradient: From pattern formation to growth regulation. Nat Rev Genet 8:663–674 - PubMed
-
- Agrawal N, Joshi S, Kango M, Saha D, Mishra A, Sinha P 1995. Epithelial hyperplasia of imaginal discs induced by mutations in Drosophila tumor suppressor genes: Growth and pattern formation in genetic mosaics. Dev Biol 169:387–398 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases