

Cytoskeletal mechanisms for breaking cellular symmetry
- PMID: 20182610
- PMCID: PMC2827899
- DOI: 10.1101/cshperspect.a003392
Cytoskeletal mechanisms for breaking cellular symmetry
Abstract
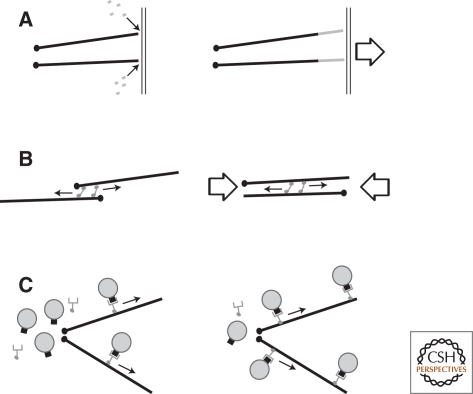
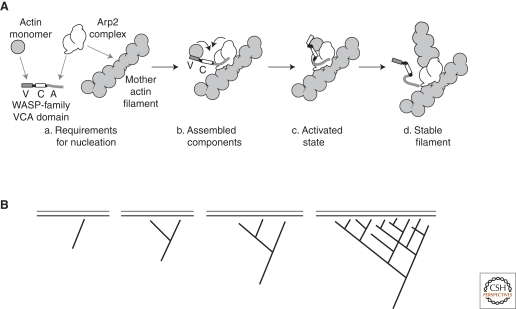
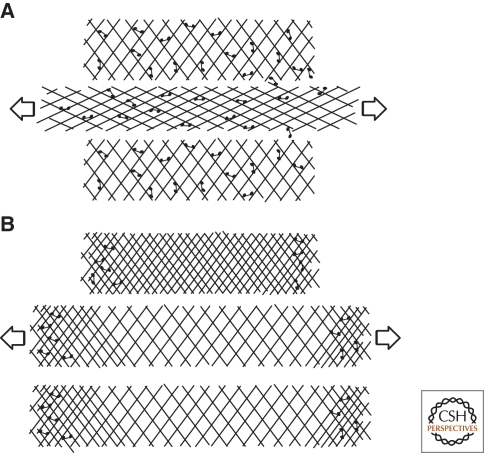
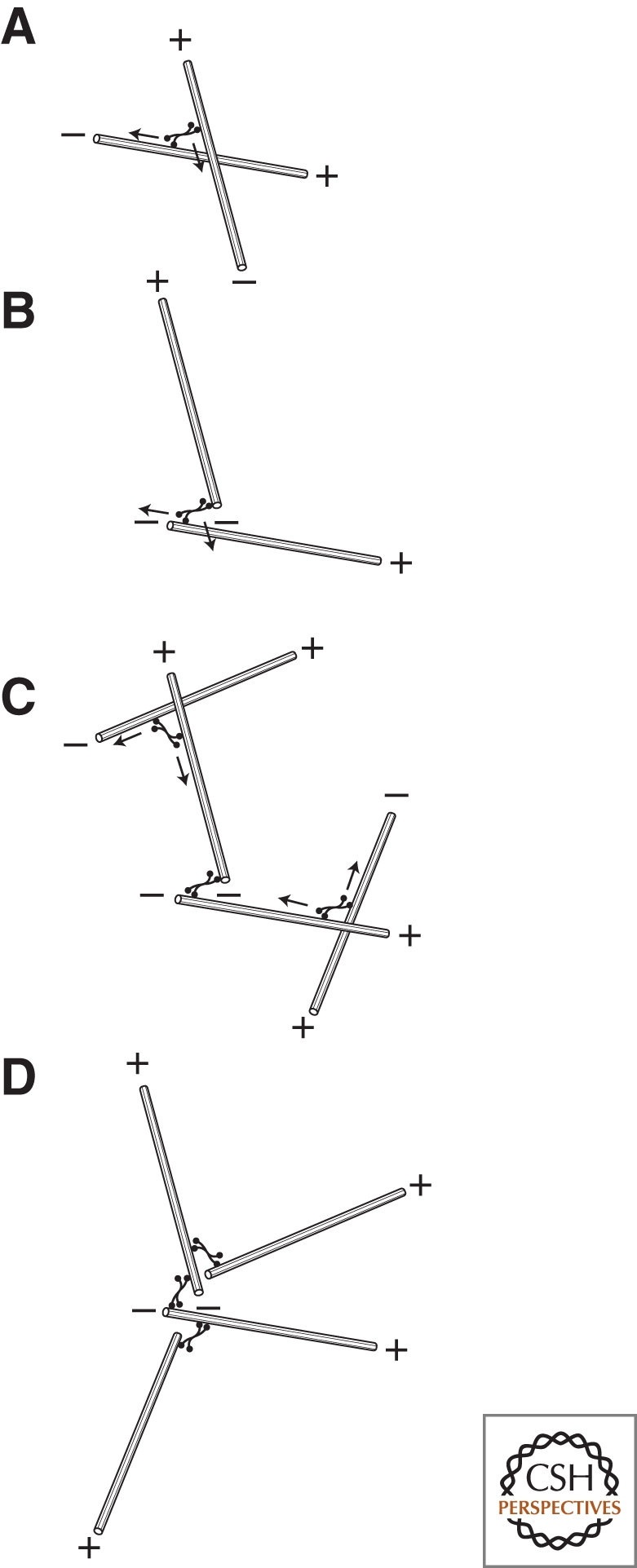
Cytoskeletal systems are networks of polymers found in all eukaryotic and many prokaryotic cells. Their purpose is to transmit and integrate information across cellular dimensions and help turn a disorderly mob of macromolecules into a spatially organized, living cell. Information, in this context, includes physical and chemical properties relevant to cellular physiology, including: the number and activity of macromolecules, cell shape, and mechanical force. Most animal cells are 10-50 microns in diameter, whereas the macromolecules that comprise them are 10,000-fold smaller (2-20 nm). To establish long-range order over cellular length scales, individual molecules must, therefore, self-assemble into larger polymers, with lengths (0.1-20 m) comparable to the size of a cell. These polymers must then be cross-linked into organized networks that fill the cytoplasm. Such cell-spanning polymer networks enable different parts of the cytoplasm to communicate directly with each other, either by transmitting forces or by carrying cargo from one spot to another.
Figures

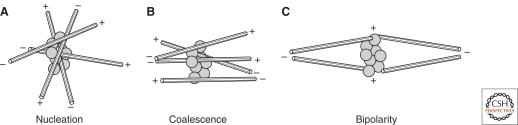
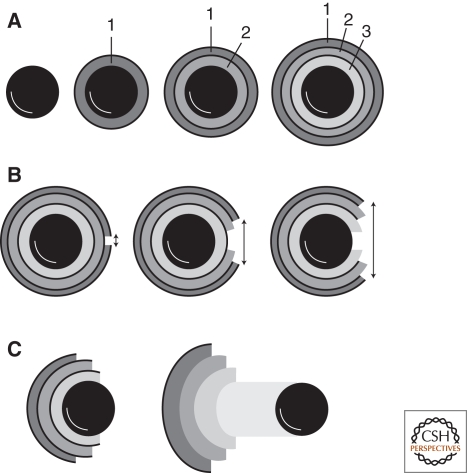
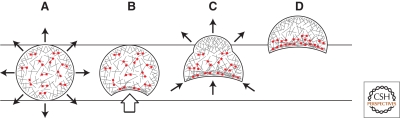
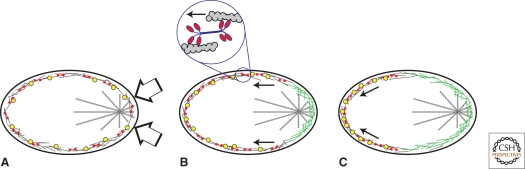
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources