

Peroxynitrite induces destruction of the tetrahydrobiopterin and heme in endothelial nitric oxide synthase: transition from reversible to irreversible enzyme inhibition
- PMID: 20184376
- PMCID: PMC2851177
- DOI: 10.1021/bi9016632
Peroxynitrite induces destruction of the tetrahydrobiopterin and heme in endothelial nitric oxide synthase: transition from reversible to irreversible enzyme inhibition
Abstract
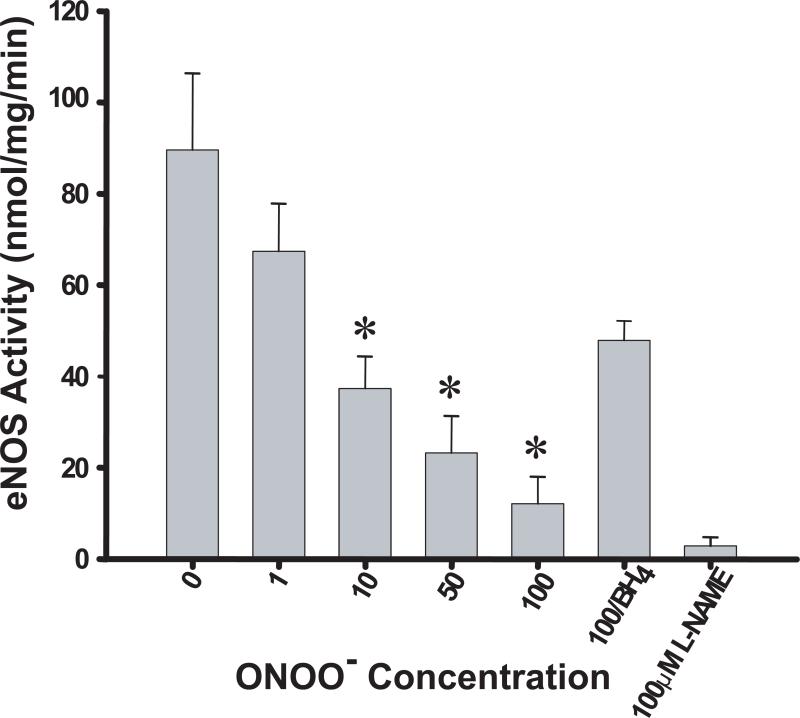
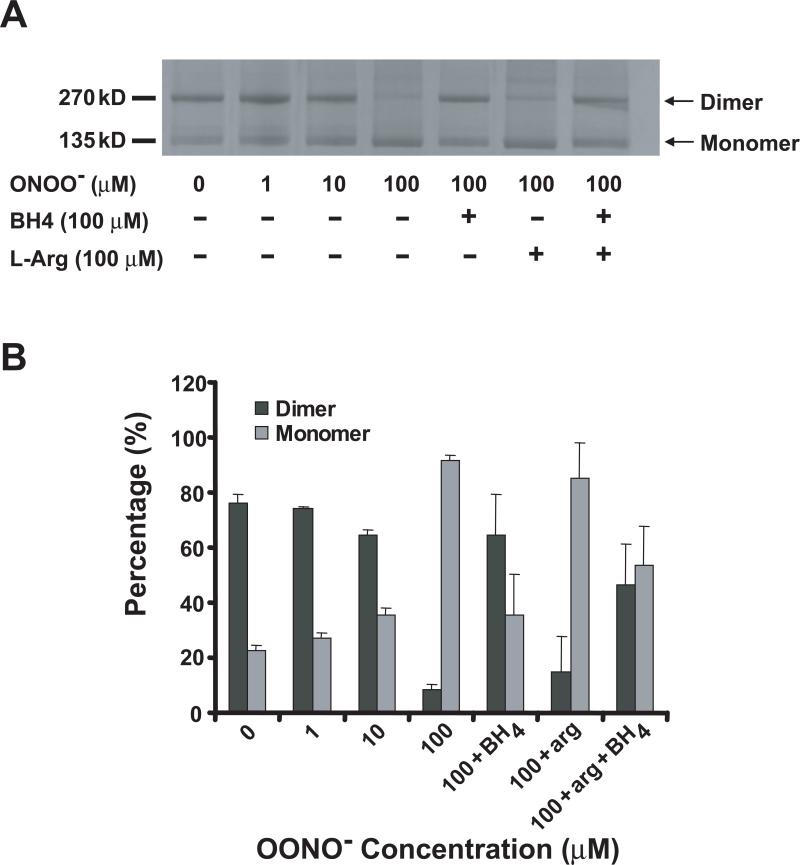
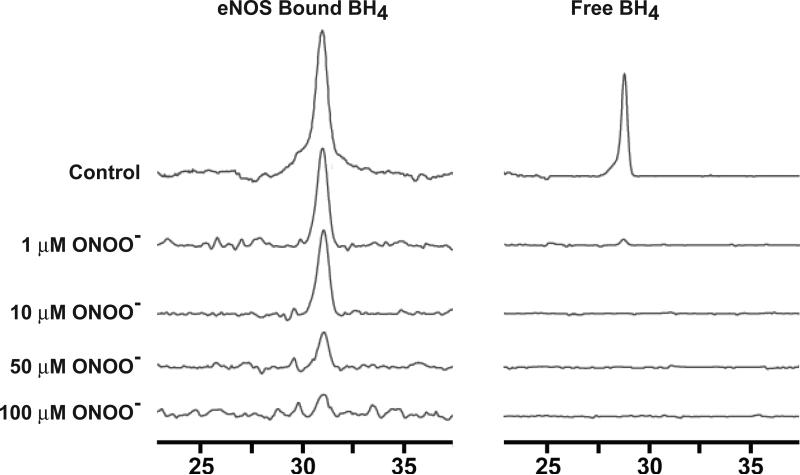
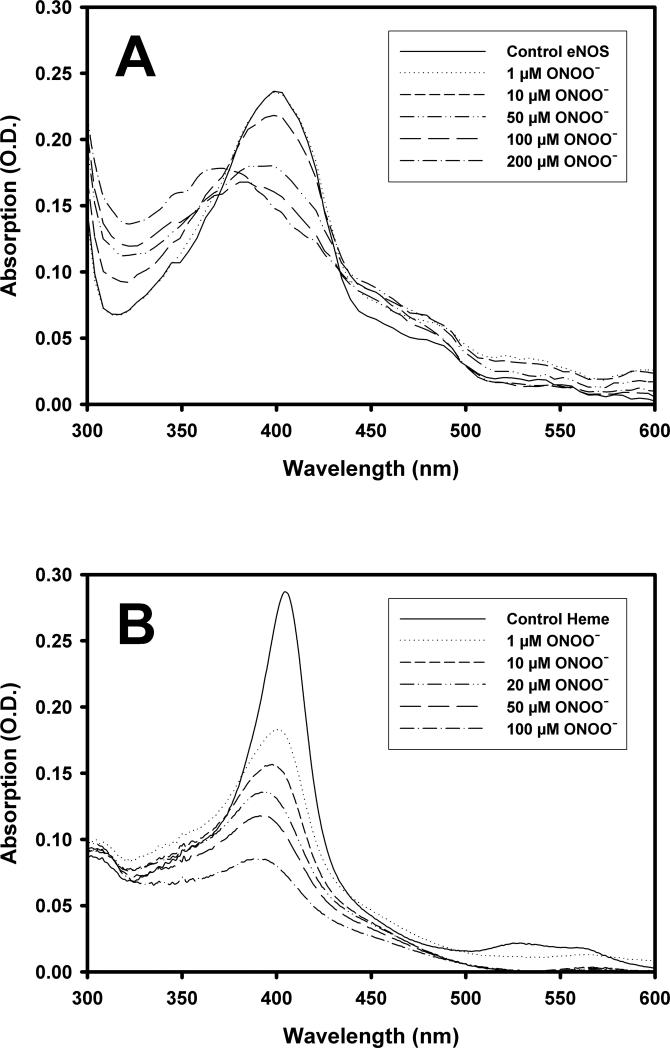
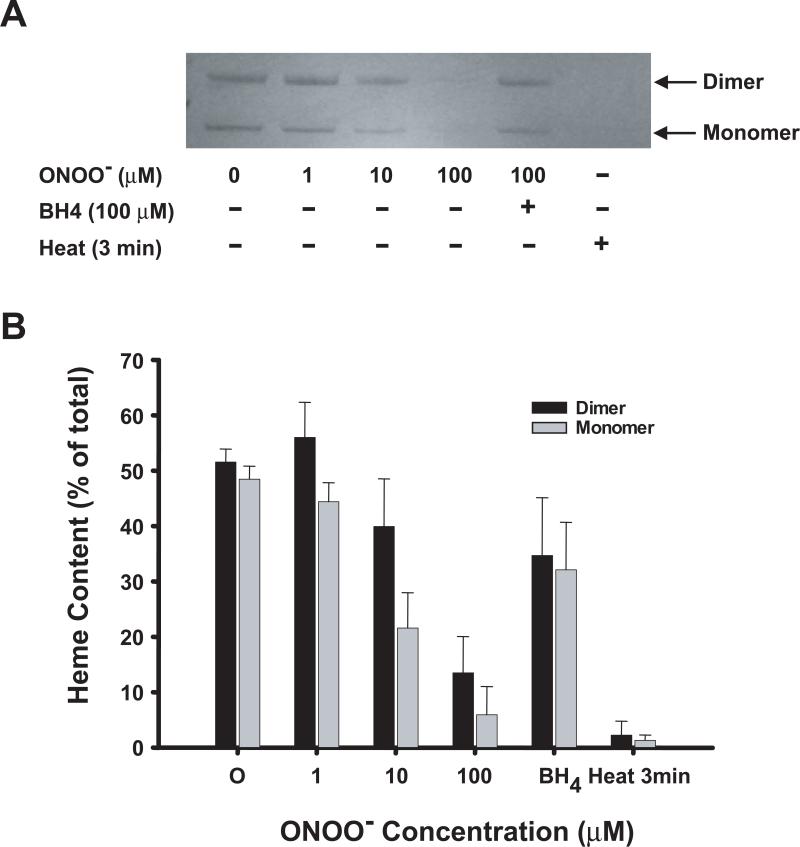
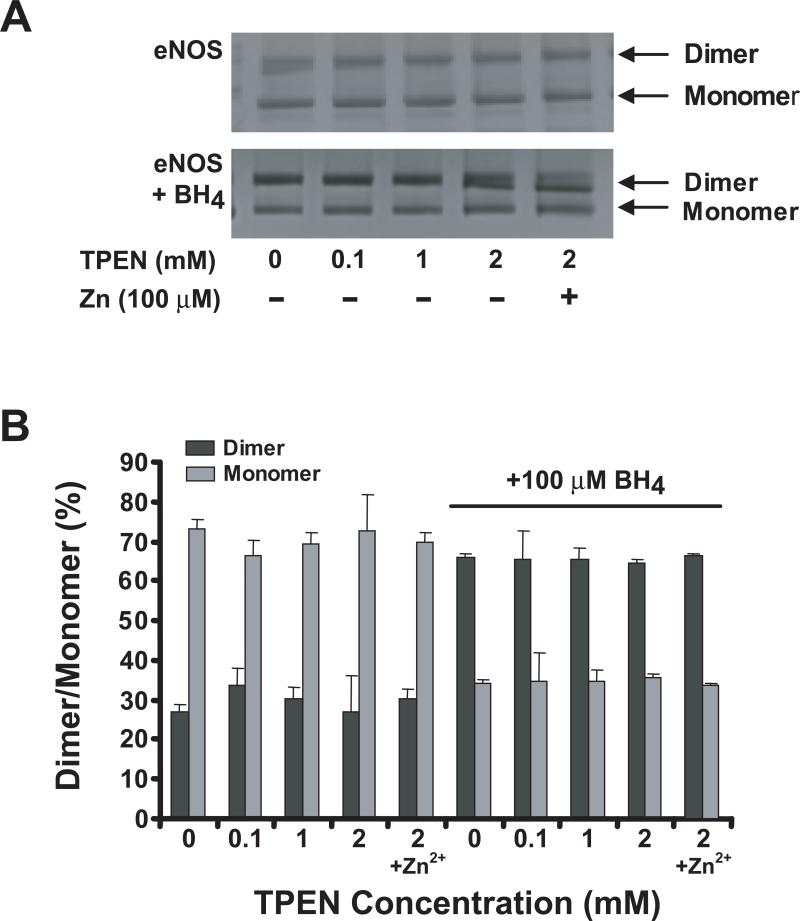
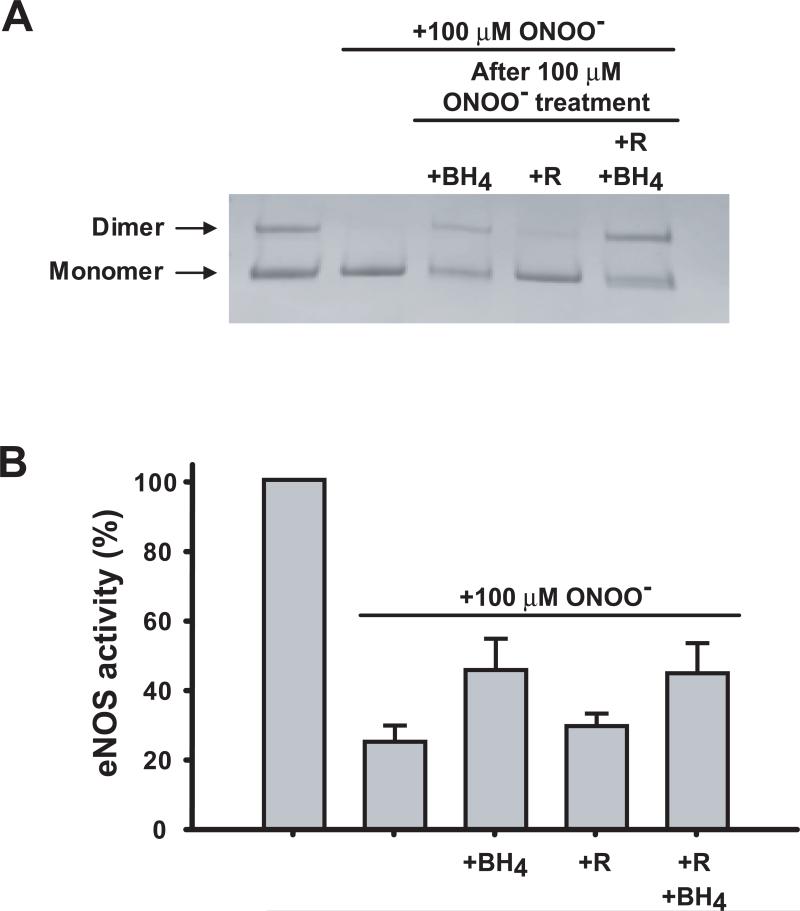
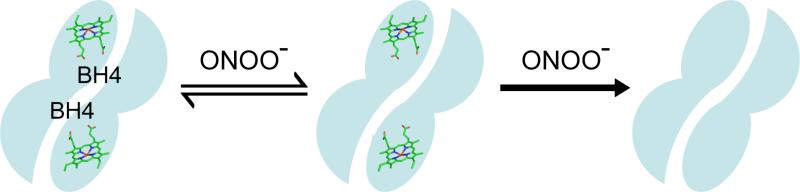
Endothelial nitric oxide synthase (eNOS) is an important regulator of vascular and cardiac function. Peroxynitrite (ONOO(-)) inactivates eNOS, but questions remain regarding the mechanisms of this process. It has been reported that inactivation is due to oxidation of the eNOS zinc-thiolate cluster, rather than the cofactor tetrahydrobiopterin (BH(4)); however, this remains highly controversial. Therefore, we investigated the mechanisms of ONOO(-)-induced eNOS dysfunction and their dose dependence. Exposure of human eNOS to ONOO(-) resulted in a dose-dependent loss of activity with a marked destabilization of the eNOS dimer. HPLC analysis indicated that both free and eNOS-bound BH(4) were oxidized during exposure to ONOO(-); however, full oxidation of protein-bound biopterin required higher ONOO(-) levels. Additionally, ONOO(-) triggered changes in the UV/visible spectrum and heme content of the enzyme. Preincubation of eNOS with BH(4) decreased dimer destabilization and heme alteration. Addition of BH(4) to the ONOO(-)-destabilized eNOS dimer only partially rescued enzyme function. In contrast to ONOO(-) treatment, incubation with the zinc chelator TPEN with removal of enzyme-bound zinc did not change the eNOS activity or stability of the SDS-resistant eNOS dimer, demonstrating that the dimer stabilization induced by BH(4) does not require zinc occupancy of the zinc-thiolate cluster. While ONOO(-) treatment was observed to induce loss of Zn binding, this cannot account for the loss of enzyme activity. Therefore, ONOO(-)-induced eNOS inactivation is primarily due to oxidation of BH(4) and irreversible destruction of the heme/heme center.
Figures
References
-
- Gross SS, Wolin MS. Nitric oxide: pathophysiological mechanisms. Annu Rev Physiol. 1995;57:737–769. - PubMed
-
- Huang PL. Endothelial nitric oxide synthase and endothelial dysfunction. Curr Hypertens Rep. 2003;5:473–480. - PubMed
-
- Fulton D, Gratton JP, Sessa WC. Post-translational control of endothelial nitric oxide synthase: why isn't calcium/calmodulin enough? J Pharmacol Exp Ther. 2001;299:818–824. - PubMed
-
- García-Cardeña G, Fan R, Shah V, Sorrentino R, Cirino G, Papapetropoulos A, Sessa WC. Dynamic activation of endothelial nitric oxide synthase by Hsp90. Nature. 1998;392:821–824. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
