

Insulation of a synthetic hydrogen metabolism circuit in bacteria
- PMID: 20184755
- PMCID: PMC2847965
- DOI: 10.1186/1754-1611-4-3
Insulation of a synthetic hydrogen metabolism circuit in bacteria
Abstract
Background: The engineering of metabolism holds tremendous promise for the production of desirable metabolites, particularly alternative fuels and other highly reduced molecules. Engineering approaches must redirect the transfer of chemical reducing equivalents, preventing these electrons from being lost to general cellular metabolism. This is especially the case for high energy electrons stored in iron-sulfur clusters within proteins, which are readily transferred when two such clusters are brought in close proximity. Iron sulfur proteins therefore require mechanisms to ensure interaction between proper partners, analogous to many signal transduction proteins. While there has been progress in the isolation of engineered metabolic pathways in recent years, the design of insulated electron metabolism circuits in vivo has not been pursued.
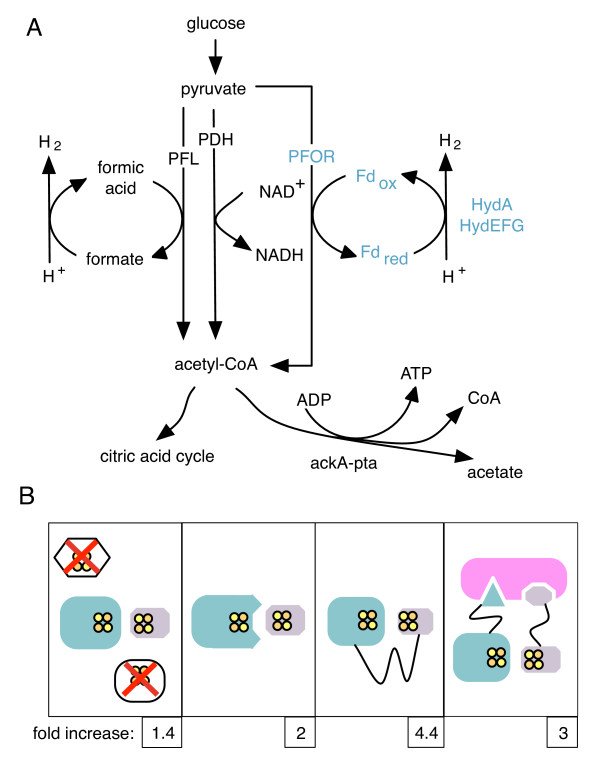
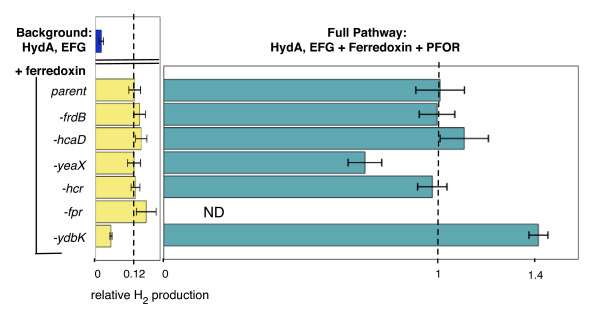
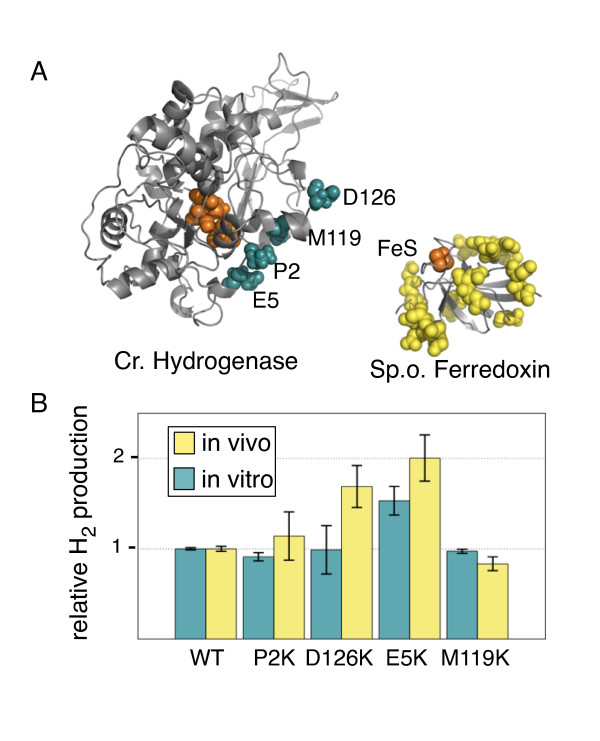
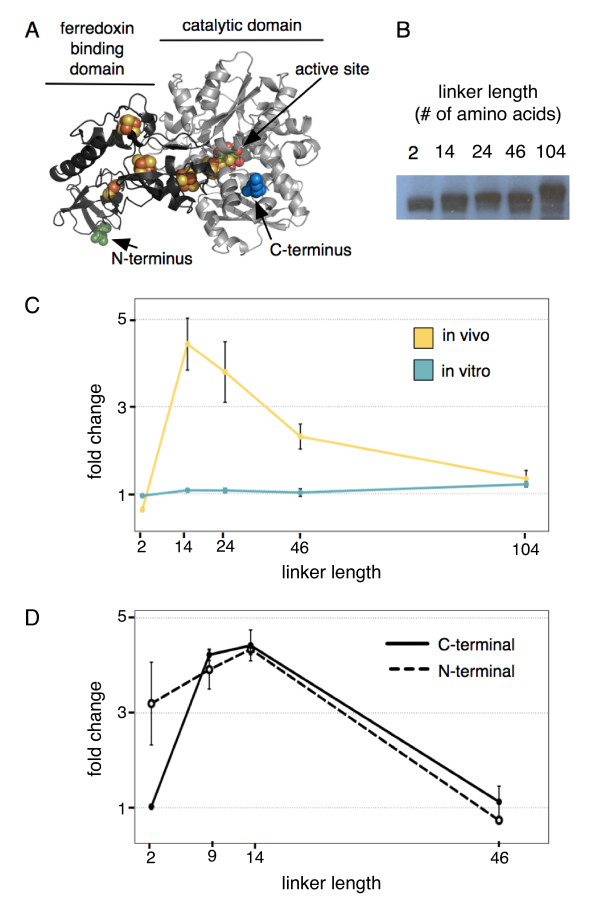
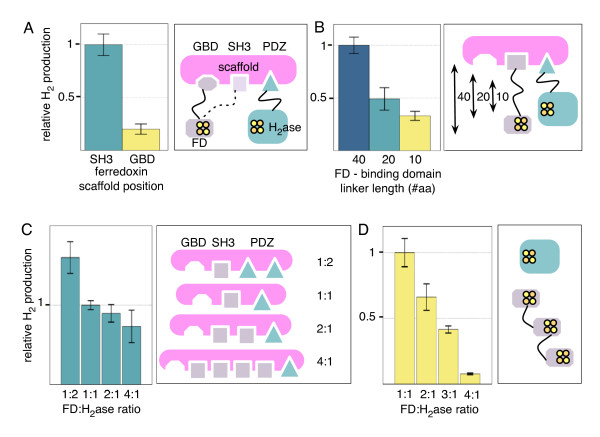
Results: Here we show that a synthetic hydrogen-producing electron transfer circuit in Escherichia coli can be insulated from existing cellular metabolism via multiple approaches, in many cases improving the function of the pathway. Our circuit is composed of heterologously expressed [Fe-Fe]-hydrogenase, ferredoxin, and pyruvate-ferredoxin oxidoreductase (PFOR), allowing the production of hydrogen gas to be coupled to the breakdown of glucose. We show that this synthetic pathway can be insulated through the deletion of competing reactions, rational engineering of protein interaction surfaces, direct protein fusion of interacting partners, and co-localization of pathway components on heterologous protein scaffolds.
Conclusions: Through the construction and characterization of a synthetic metabolic circuit in vivo, we demonstrate a novel system that allows for predictable engineering of an insulated electron transfer pathway. The development of this system demonstrates working principles for the optimization of engineered pathways for alternative energy production, as well as for understanding how electron transfer between proteins is controlled.
Figures

Similar articles
-
Modular electron transfer circuits for synthetic biology: insulation of an engineered biohydrogen pathway.Bioeng Bugs. 2010 Nov-Dec;1(6):413-8. doi: 10.4161/bbug.1.6.12462. Bioeng Bugs. 2010. PMID: 21468209 Free PMC article.
-
Expanding the substrates for a bacterial hydrogenlyase reaction.Microbiology (Reading). 2017 May;163(5):649-653. doi: 10.1099/mic.0.000471. Microbiology (Reading). 2017. PMID: 28488566 Free PMC article.
-
Defining optimal electron transfer partners for light-driven cytochrome P450 reactions.Metab Eng. 2019 Sep;55:33-43. doi: 10.1016/j.ymben.2019.05.003. Epub 2019 May 12. Metab Eng. 2019. PMID: 31091467
-
Photosynthetic fuel for heterologous enzymes: the role of electron carrier proteins.Photosynth Res. 2017 Dec;134(3):329-342. doi: 10.1007/s11120-017-0364-0. Epub 2017 Mar 11. Photosynth Res. 2017. PMID: 28285375 Review.
-
Structural principles for computational and de novo design of 4Fe-4S metalloproteins.Biochim Biophys Acta. 2016 May;1857(5):531-538. doi: 10.1016/j.bbabio.2015.10.001. Epub 2015 Oct 9. Biochim Biophys Acta. 2016. PMID: 26449207 Free PMC article. Review.
Cited by
-
Designed surface residue substitutions in [NiFe] hydrogenase that improve electron transfer characteristics.Int J Mol Sci. 2015 Jan 16;16(1):2020-33. doi: 10.3390/ijms16012020. Int J Mol Sci. 2015. PMID: 25603181 Free PMC article.
-
Membrane-Located Expression of Thioesterase From Acinetobacter baylyi Enhances Free Fatty Acid Production With Decreased Toxicity in Synechocystis sp. PCC6803.Front Microbiol. 2018 Nov 27;9:2842. doi: 10.3389/fmicb.2018.02842. eCollection 2018. Front Microbiol. 2018. PMID: 30538684 Free PMC article.
-
Hydrogen production by recombinant Escherichia coli strains.Microb Biotechnol. 2012 Mar;5(2):214-25. doi: 10.1111/j.1751-7915.2011.00282.x. Epub 2011 Sep 6. Microb Biotechnol. 2012. PMID: 21895995 Free PMC article. Review.
-
Research progress and the biotechnological applications of multienzyme complex.Appl Microbiol Biotechnol. 2021 Mar;105(5):1759-1777. doi: 10.1007/s00253-021-11121-4. Epub 2021 Feb 10. Appl Microbiol Biotechnol. 2021. PMID: 33564922 Review.
-
Theoretical hypothesis in a direct electron transfer between non-interacting Fe-S proteins within an artificial fusion.FEMS Microbiol Lett. 2024 Jan 9;371:fnad137. doi: 10.1093/femsle/fnad137. FEMS Microbiol Lett. 2024. PMID: 38196139 Free PMC article.
References
-
- Marcus RA, Sutin N. Electron transfers in chemistry and biology. Biochim Biophys Acta. 1985;811:265–322.
-
- Gou P, Hanke GT, Kimata-Ariga Y, Standley DM, Kubo A, Taniguchi I, Nakamura H, Hase T. Higher order structure contributes to specific differences in redox potential and electron transfer efficiency of root and leaf ferredoxins. Biochemistry. 2006;45:14389–14396. doi: 10.1021/bi061779d. - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
