

Initiation of DNA replication after fertilization is regulated by p90Rsk at pre-RC/pre-IC transition in starfish eggs
- PMID: 20185755
- PMCID: PMC2841915
- DOI: 10.1073/pnas.1000587107
Initiation of DNA replication after fertilization is regulated by p90Rsk at pre-RC/pre-IC transition in starfish eggs
Abstract
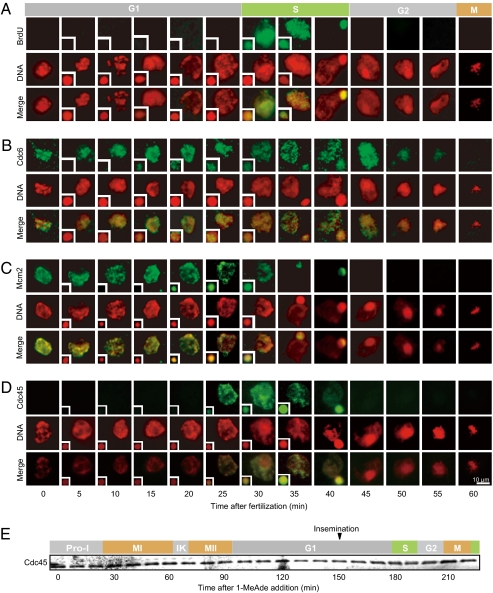
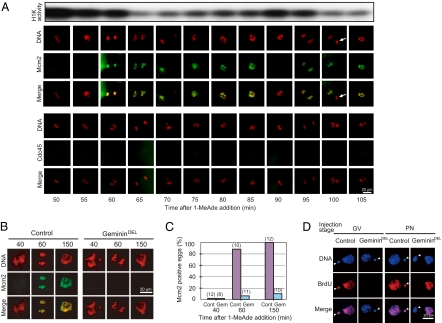
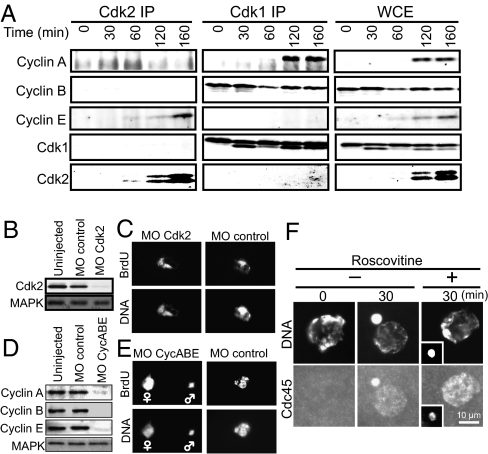
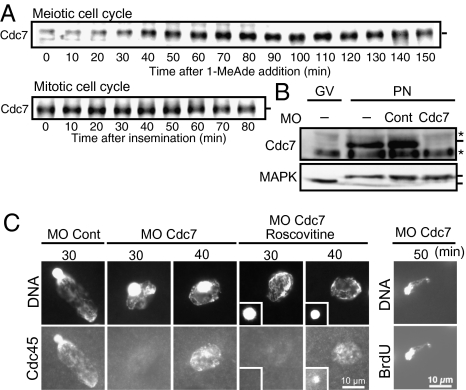
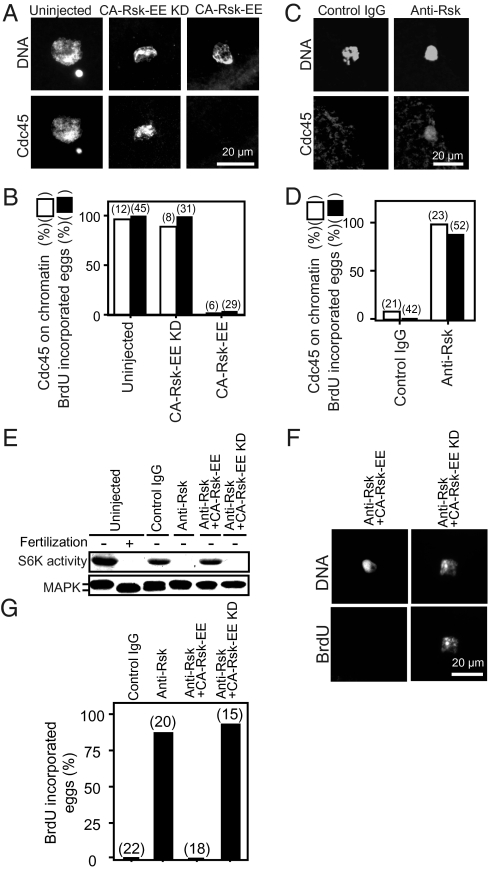
Initiation of DNA replication in eukaryotic cells is controlled through an ordered assembly of protein complexes at replication origins. The molecules involved in this process are well conserved but diversely regulated. Typically, initiation of DNA replication is regulated in response to developmental events in multicellular organisms. Here, we elucidate the regulation of the first S phase of the embryonic cell cycle after fertilization. Unless fertilization occurs, the Mos-MAPK-p90Rsk pathway causes the G1-phase arrest after completion of meiosis in starfish eggs. Fertilization shuts down this pathway, leading to the first S phase with no requirement of new protein synthesis. However, how and in which stage the initiation complex for DNA replication is arrested by p90Rsk remains unclear. We find that in G1-arrested eggs, chromatin is loaded with the Mcm complex to form the prereplicative complex (pre-RC). Inactivation of p90Rsk is necessary and sufficient for further loading of Cdc45 onto chromatin to form the preinitiation complex (pre-IC) and the subsequent initiation of DNA replication. However, cyclin A-, B-, and E-Cdk's activity and Cdc7 accumulation are dispensable for these processes. These observations define the stage of G1 arrest in unfertilized eggs at transition point from pre-RC to pre-IC, and reveal a unique role of p90Rsk for a negative regulator of this transition. Thus, initiation of DNA replication in the meiosis-to-mitosis transition is regulated at the pre-RC stage as like in the G1 checkpoint, but in a manner different from the checkpoint.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Machida YJ, Hamlin JL, Dutta A. Right place, right time, and only once: Replication initiation in metazoans. Cell. 2005;123:13–24. - PubMed
-
- Kearsey SE, Cotterill S. Enigmatic variations: Divergent modes of regulating eukaryotic DNA replication. Mol Cell. 2003;12:1067–1075. - PubMed
-
- Meijer L, Guerrier P. Maturation and fertilization in starfish oocytes. Int Rev Cytol. 1984;86:129–196. - PubMed
-
- Page AW, Orr-Weaver TL. Stopping and starting the meiotic cell cycle. Curr Opin Genet Dev. 1997;7:23–31. - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
