

Glucose intolerance and reduced proliferation of pancreatic beta-cells in transgenic pigs with impaired glucose-dependent insulinotropic polypeptide function
- PMID: 20185813
- PMCID: PMC2857903
- DOI: 10.2337/db09-0519
Glucose intolerance and reduced proliferation of pancreatic beta-cells in transgenic pigs with impaired glucose-dependent insulinotropic polypeptide function
Abstract
Objective: The insulinotropic action of the incretin glucose-dependent insulinotropic polypeptide (GIP) is impaired in type 2 diabetes, while the effect of glucagon-like peptide-1 (GLP-1) is preserved. To evaluate the role of impaired GIP function in glucose homeostasis and development of the endocrine pancreas in a large animal model, we generated transgenic pigs expressing a dominant-negative GIP receptor (GIPR(dn)) in pancreatic islets.
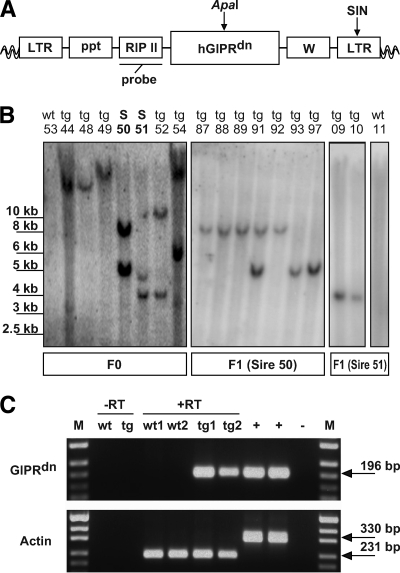
Research design and methods: GIPR(dn) transgenic pigs were generated using lentiviral transgenesis. Metabolic tests and quantitative stereological analyses of the different endocrine islet cell populations were performed, and beta-cell proliferation and apoptosis were quantified to characterize this novel animal model.
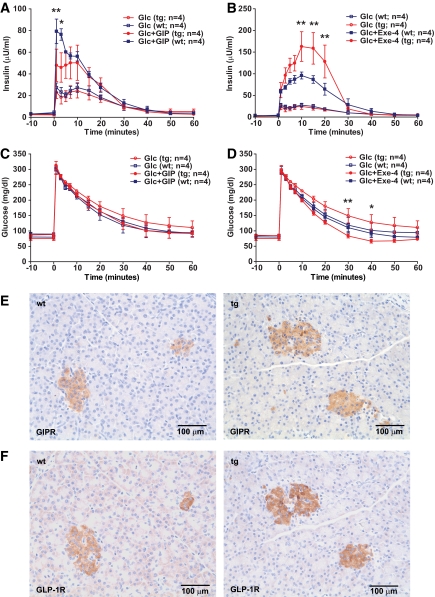
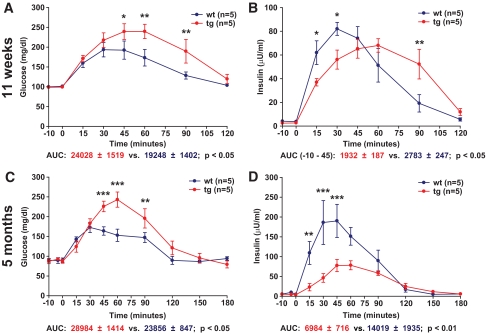
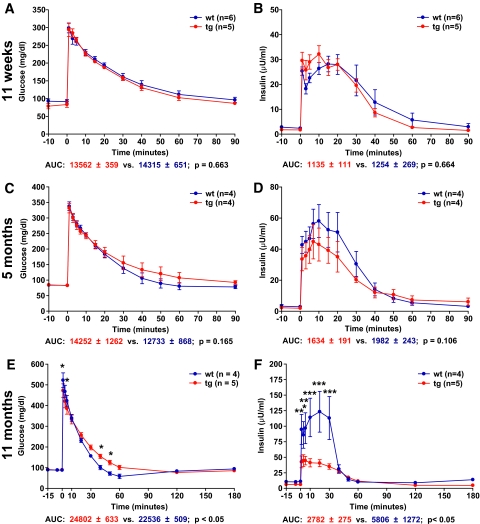
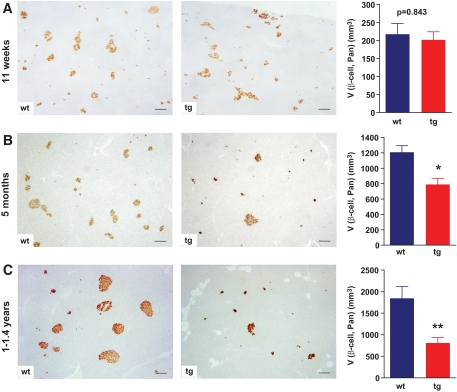
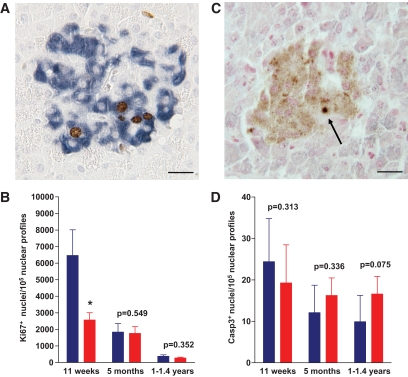
Results: Eleven-week-old GIPR(dn) transgenic pigs exhibited significantly reduced oral glucose tolerance due to delayed insulin secretion, whereas intravenous glucose tolerance and pancreatic beta-cell mass were not different from controls. The insulinotropic effect of GIP was significantly reduced, whereas insulin secretion in response to the GLP-1 receptor agonist exendin-4 was enhanced in GIPR(dn) transgenic versus control pigs. With increasing age, glucose control deteriorated in GIPR(dn) transgenic pigs, as shown by reduced oral and intravenous glucose tolerance due to impaired insulin secretion. Importantly, beta-cell proliferation was reduced by 60% in 11-week-old GIPR(dn) transgenic pigs, leading to a reduction of beta-cell mass by 35% and 58% in 5-month-old and 1- to 1.4-year-old transgenic pigs compared with age-matched controls, respectively.
Conclusions: The first large animal model with impaired incretin function demonstrates an essential role of GIP for insulin secretion, proliferation of beta-cells, and physiological expansion of beta-cell mass.
Figures
References
-
- Baggio LL, Drucker DJ. Biology of incretins: GLP-1 and GIP. Gastroenterology 2007;132:2131–2157 - PubMed
-
- Mayo KE, Miller LJ, Bataille D, Dalle S, Goke B, Thorens B, Drucker DJ. International Union of Pharmacology. XXXV. The glucagon receptor family. Pharmacol Rev 2003;55:167–194 - PubMed
-
- Holst JJ, Gromada J. Role of incretin hormones in the regulation of insulin secretion in diabetic and nondiabetic humans. Am J Physiol Endocrinol Metab 2004;287:E199–E206 - PubMed
-
- Saxena R, Hivert MF, Langenberg C, Tanaka T, Pankow JS, Vollenweider P, Lyssenko V, Bouatia-Naji N, Dupuis J, Jackson AU, Kao WH, Li M, Glazer NL, Manning AK, Luan J, Stringham HM, Prokopenko I, Johnson T, Grarup N, Boesgaard TW, Lecoeur C, Shrader P, O'Connell J, Ingelsson E, Couper DJ, Rice K, Song K, Andreasen CH, Dina C, Kottgen A, Le Bacquer O, Pattou F, Taneera J, Steinthorsdottir V, Rybin D, Ardlie K, Sampson M, Qi L, van Hoek M, Weedon MN, Aulchenko YS, Voight BF, Grallert H, Balkau B, Bergman RN, Bielinski SJ, Bonnefond A, Bonnycastle LL, Borch-Johnsen K, Bottcher Y, Brunner E, Buchanan TA, Bumpstead SJ, Cavalcanti-Proenca C, Charpentier G, Chen YD, Chines PS, Collins FS, Cornelis M, G JC, Delplanque J, Doney A, Egan JM, Erdos MR, Firmann M, Forouhi NG, Fox CS, Goodarzi MO, Graessler J, Hingorani A, Isomaa B, Jorgensen T, Kivimaki M, Kovacs P, Krohn K, Kumari M, Lauritzen T, Levy-Marchal C, Mayor V, McAteer JB, Meyre D, Mitchell BD, Mohlke KL, Morken MA, Narisu N, Palmer CN, Pakyz R, Pascoe L, Payne F, Pearson D, Rathmann W, Sandbaek A, Sayer AA, Scott LJ, Sharp SJ, Sijbrands E, Singleton A, Siscovick DS, Smith NL, Sparso T, Swift AJ, Syddall H, Thorleifsson G, Tonjes A, Tuomi T, Tuomilehto J, Valle TT, Waeber G, Walley A, Waterworth DM, Zeggini E, Zhao JH, Illig T, Wichmann HE, Wilson JF, van Duijn C, Hu FB, Morris AD, Frayling TM, Hattersley AT, Thorsteinsdottir U, Stefansson K, Nilsson P, Syvanen AC, Shuldiner AR, Walker M, Bornstein SR, Schwarz P, Williams GH, Nathan DM, Kuusisto J, Laakso M, Cooper C, Marmot M, Ferrucci L, Mooser V, Stumvoll M, Loos RJ, Altshuler D, Psaty BM, Rotter JI, Boerwinkle E, Hansen T, Pedersen O, Florez JC, McCarthy MI, Boehnke M, Barroso I, Sladek R, Froguel P, Meigs JB, Groop L, Wareham NJ, Watanabe RM: Genetic variation in GIPR influences the glucose and insulin responses to an oral glucose challenge. Nat Genet 2010;42:142–148 - PMC - PubMed
-
- Ehses JA, Casilla VR, Doty T, Pospisilik JA, Winter KD, Demuth HU, Pederson RA, McIntosh CH. Glucose-dependent insulinotropic polypeptide promotes beta-(INS-1) cell survival via cyclic adenosine monophosphate-mediated caspase-3 inhibition and regulation of p38 mitogen-activated protein kinase. Endocrinology 2003;144:4433–4445 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
