

Activation of acid-sensing ion channel 1a (ASIC1a) by surface trafficking
- PMID: 20185828
- PMCID: PMC2857104
- DOI: 10.1074/jbc.M109.086041
Activation of acid-sensing ion channel 1a (ASIC1a) by surface trafficking
Abstract
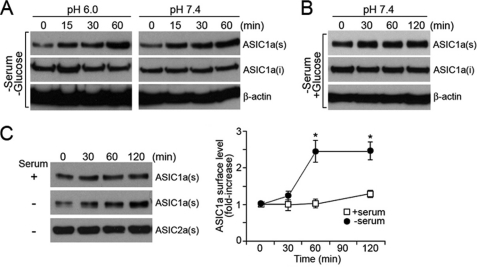
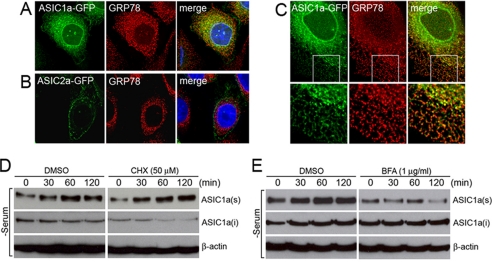
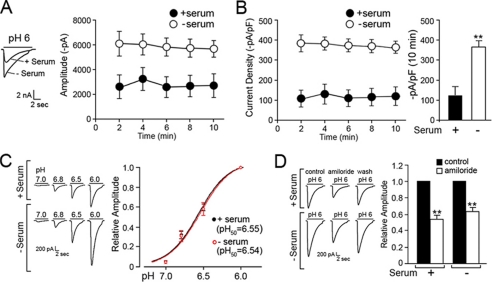
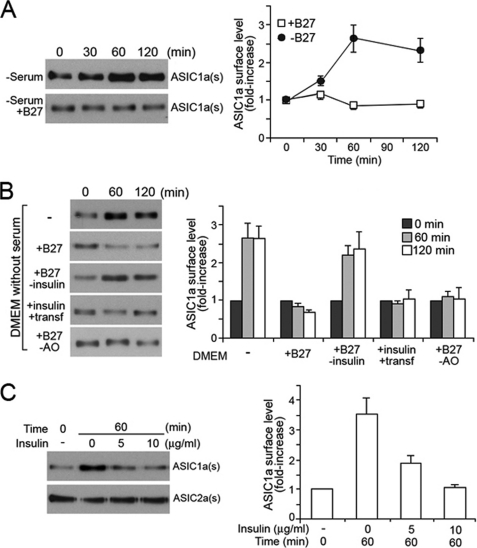
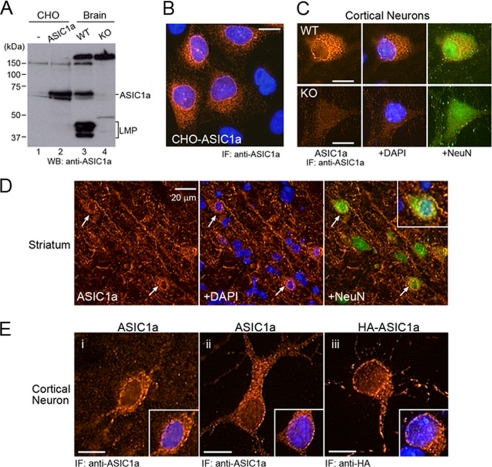
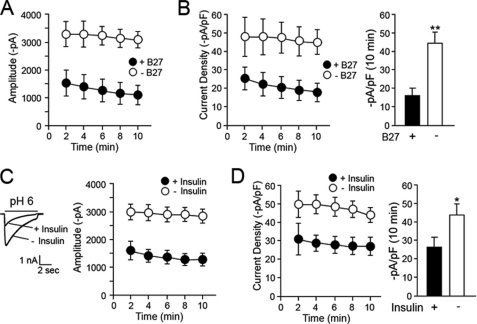
Acid-sensing ion channels (ASICs) are voltage-independent Na(+) channels activated by extracellular protons. ASIC1a is expressed in neurons in mammalian brain and is implicated in long term potentiation of synaptic transmission that contributes to learning and memory. In ischemic brain injury, however, activation of this Ca(2+)-permeable channel plays a critical role in acidosis-mediated, glutamate-independent, Ca(2+) toxicity. We report here the identification of insulin as a regulator of ASIC1a surface expression. In modeled ischemia using Chinese hamster ovary cells, serum depletion caused a significant increase in ASIC1a surface expression that resulted in the potentiation of ASIC1a activity. Among the components of serum, insulin was identified as the key factor that maintains a low level of ASIC1a on the plasma membrane. Neurons subjected to insulin depletion increased surface expression of ASIC1a with resultant potentiation of ASIC1a currents. Intracellularly, ASIC1a is predominantly localized to the endoplasmic reticulum in Chinese hamster ovary cells, and this intracellular localization is also observed in neurons. Under conditions of serum or insulin depletion, the intracellular ASIC1a is translocated to the cell surface, increasing the surface expression level. These results reveal an important trafficking mechanism of ASIC1a that is relevant to both the normal physiology and the pathological activity of this channel.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
