

Myelin activates FAK/Akt/NF-kappaB pathways and provokes CR3-dependent inflammatory response in murine system
- PMID: 20186338
- PMCID: PMC2826415
- DOI: 10.1371/journal.pone.0009380
Myelin activates FAK/Akt/NF-kappaB pathways and provokes CR3-dependent inflammatory response in murine system
Abstract
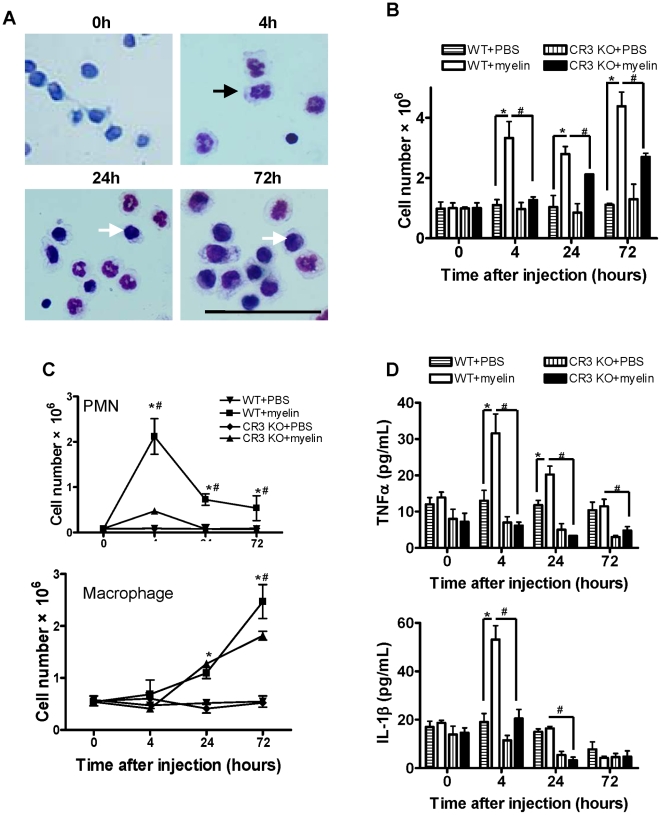
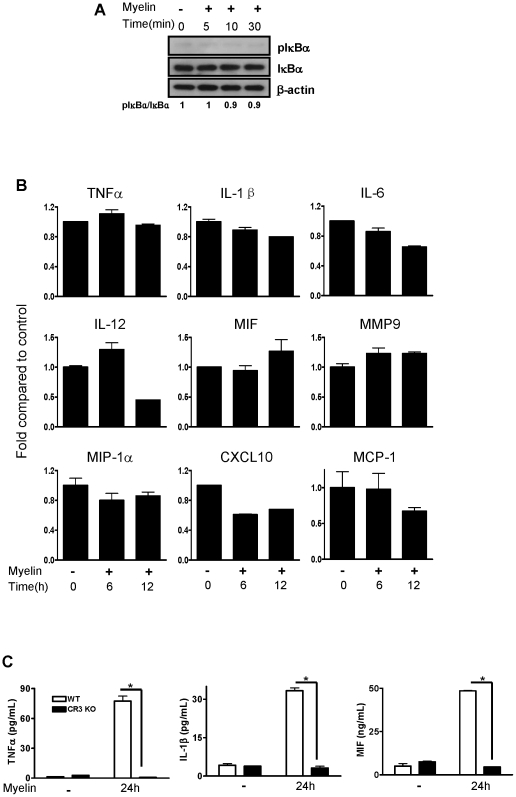
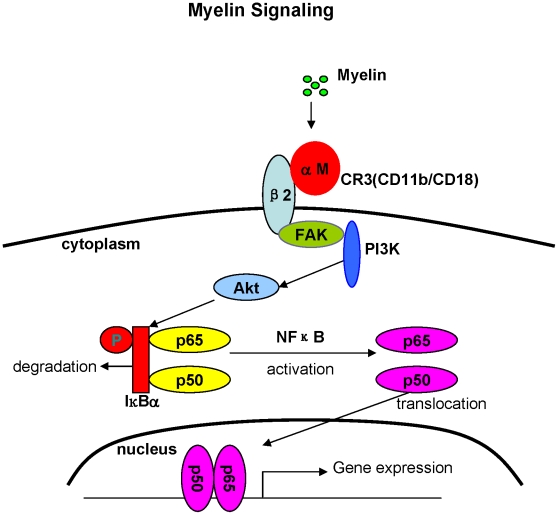
Inflammatory response following central nervous system (CNS) injury contributes to progressive neuropathology and reduction in functional recovery. Axons are sensitive to mechanical injury and toxic inflammatory mediators, which may lead to demyelination. Although it is well documented that degenerated myelin triggers undesirable inflammatory responses in autoimmune diseases such as multiple sclerosis (MS) and its animal model, experimental autoimmune encephalomyelitis (EAE), there has been very little study of the direct inflammatory consequences of damaged myelin in spinal cord injury (SCI), i.e., there is no direct evidence to show that myelin debris from injured spinal cord can trigger undesirable inflammation in vitro and in vivo. Our data showed that myelin can initiate inflammatory responses in vivo, which is complement receptor 3 (CR3)-dependent via stimulating macrophages to express pro-inflammatory molecules and down-regulates expression of anti-inflammatory cytokines. Mechanism study revealed that myelin-increased cytokine expression is through activation of FAK/PI3K/Akt/NF-kappaB signaling pathways and CR3 contributes to myelin-induced PI3K/Akt/NF-kappaB activation and cytokine production. The myelin induced inflammatory response is myelin specific as sphingomyelin (the major lipid of myelin) and myelin basic protein (MBP, one of the major proteins of myelin) are not able to activate NF-kappaB signaling pathway. In conclusion, our results demonstrate a crucial role of myelin as an endogenous inflammatory stimulus that induces pro-inflammatory responses and suggest that blocking myelin-CR3 interaction and enhancing myelin debris clearance may be effective interventions for treating SCI.
Conflict of interest statement
Figures







References
-
- Hausmann ON. Post-traumatic inflammation following spinal cord injury. Spinal Cord. 2003;41:369–378. - PubMed
-
- Popovich PG, Wei P, Stokes BT. Cellular inflammatory response after spinal cord injury in Sprague-Dawley and Lewis rats. J Comp Neurol. 1997;377:443–464. - PubMed
-
- Pineau I, Lacroix S. Proinflammatory cytokine synthesis in the injured mouse spinal cord: multiphasic expression pattern and identification of the cell types involved. J Comp Neurol. 2007;500:267–285. - PubMed
-
- Profyris C, Cheema SS, Zang D, Azari MF, Boyle K, et al. Degenerative and regenerative mechanisms governing spinal cord injury. Neurobiol Dis. 2004;15:415–436. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
