

Ero1alpha requires oxidizing and normoxic conditions to localize to the mitochondria-associated membrane (MAM)
- PMID: 20186508
- PMCID: PMC3006622
- DOI: 10.1007/s12192-010-0174-1
Ero1alpha requires oxidizing and normoxic conditions to localize to the mitochondria-associated membrane (MAM)
Abstract
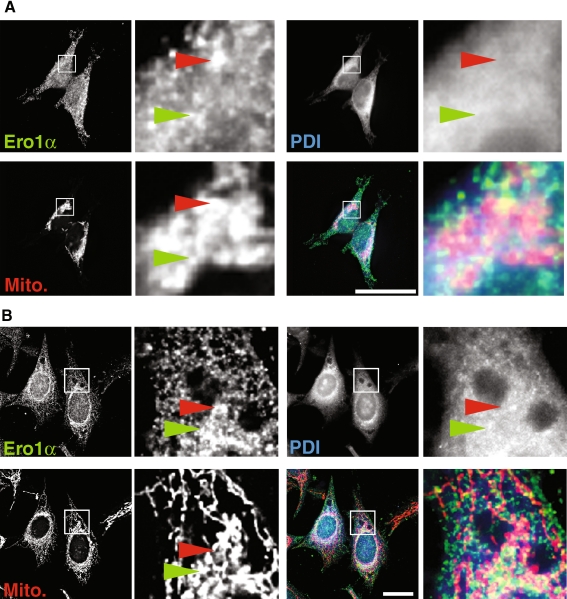
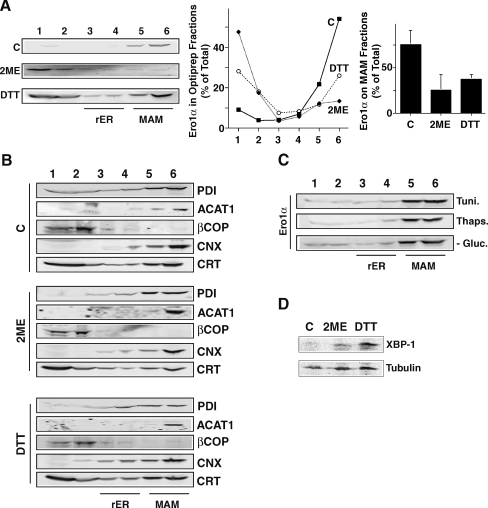
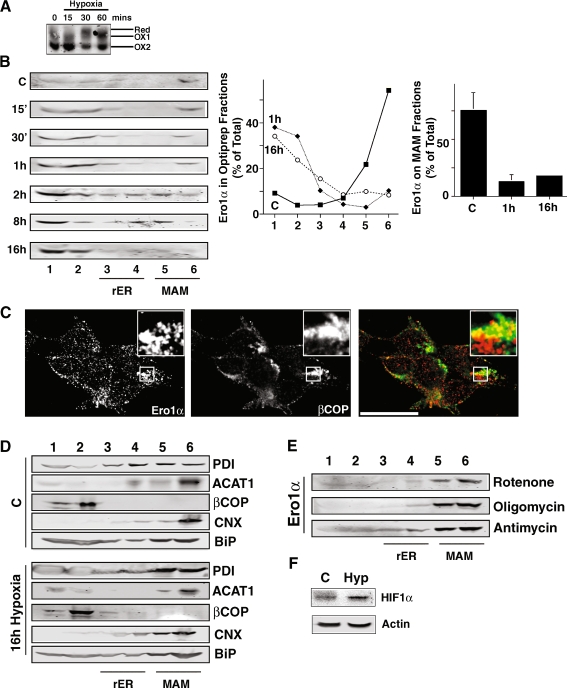
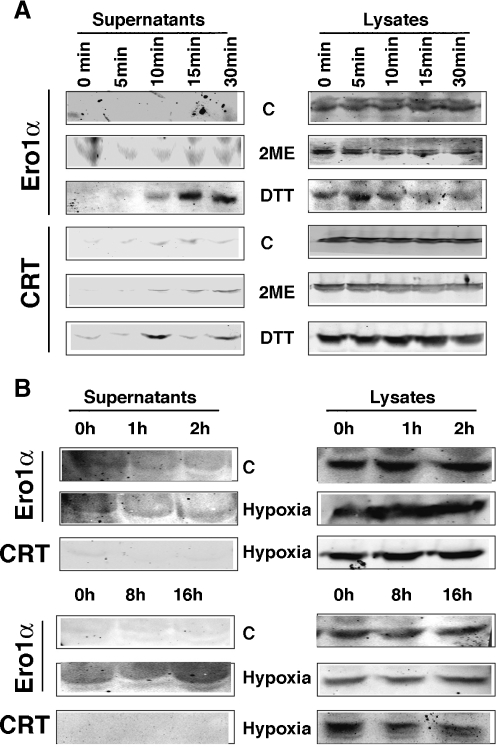
Protein secretion from the endoplasmic reticulum (ER) requires the enzymatic activity of chaperones and oxidoreductases that fold polypeptides and form disulfide bonds within newly synthesized proteins. The best-characterized ER redox relay depends on the transfer of oxidizing equivalents from molecular oxygen through ER oxidoreductin 1 (Ero1) and protein disulfide isomerase to nascent polypeptides. The formation of disulfide bonds is, however, not the sole function of ER oxidoreductases, which are also important regulators of ER calcium homeostasis. Given the role of human Ero1alpha in the regulation of the calcium release by inositol 1,4,5-trisphosphate receptors during the onset of apoptosis, we hypothesized that Ero1alpha may have a redox-sensitive localization to specific domains of the ER. Our results show that within the ER, Ero1alpha is almost exclusively found on the mitochondria-associated membrane (MAM). The localization of Ero1alpha on the MAM is dependent on oxidizing conditions within the ER. Chemical reduction of the ER environment, but not ER stress in general leads to release of Ero1alpha from the MAM. In addition, the correct localization of Ero1alpha to the MAM also requires normoxic conditions, but not ongoing oxidative phosphorylation.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
