

Knockdown of DISC1 by in utero gene transfer disturbs postnatal dopaminergic maturation in the frontal cortex and leads to adult behavioral deficits
- PMID: 20188653
- PMCID: PMC3084528
- DOI: 10.1016/j.neuron.2010.01.019
Knockdown of DISC1 by in utero gene transfer disturbs postnatal dopaminergic maturation in the frontal cortex and leads to adult behavioral deficits
Abstract
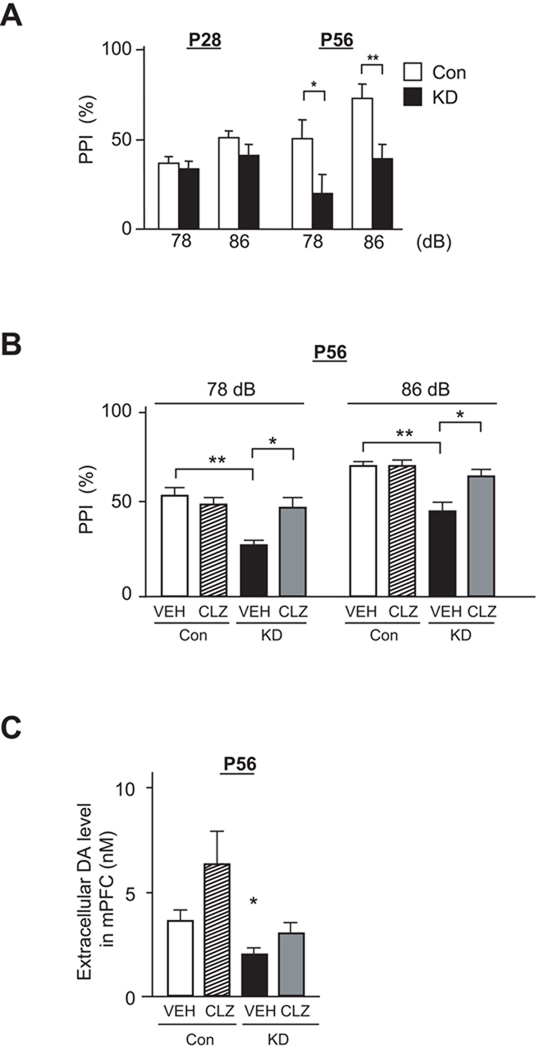
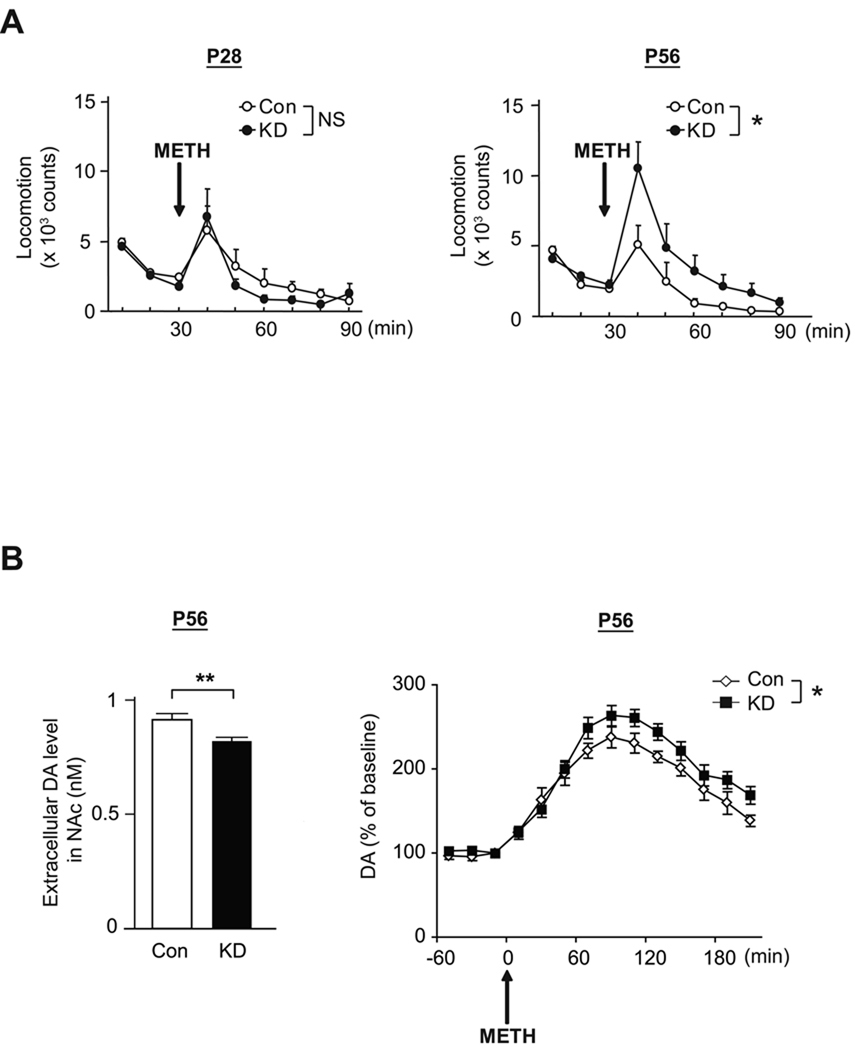
Adult brain function and behavior are influenced by neuronal network formation during development. Genetic susceptibility factors for adult psychiatric illnesses, such as Neuregulin-1 and Disrupted-in-Schizophrenia-1 (DISC1), influence adult high brain functions, including cognition and information processing. These factors have roles during neurodevelopment and are likely to cooperate, forming pathways or "signalosomes." Here we report the potential to generate an animal model via in utero gene transfer in order to address an important question of how nonlethal deficits in early development may affect postnatal brain maturation and high brain functions in adulthood, which are impaired in various psychiatric illnesses such as schizophrenia. We show that transient knockdown of DISC1 in the pre- and perinatal stages, specifically in a lineage of pyramidal neurons mainly in the prefrontal cortex, leads to selective abnormalities in postnatal mesocortical dopaminergic maturation and behavioral abnormalities associated with disturbed cortical neurocircuitry after puberty.
Figures
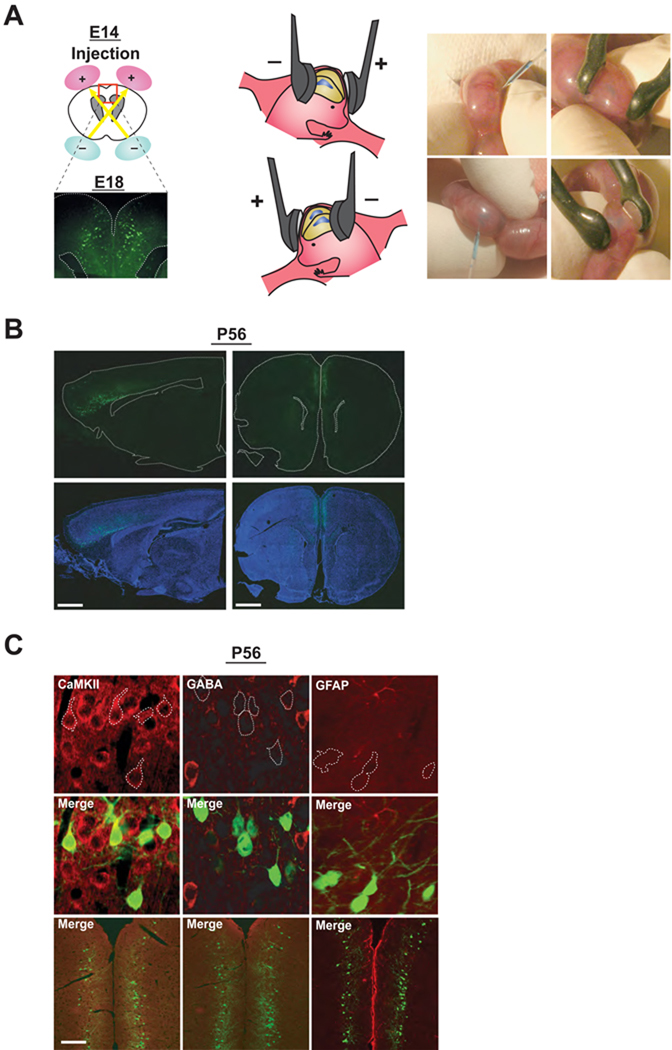
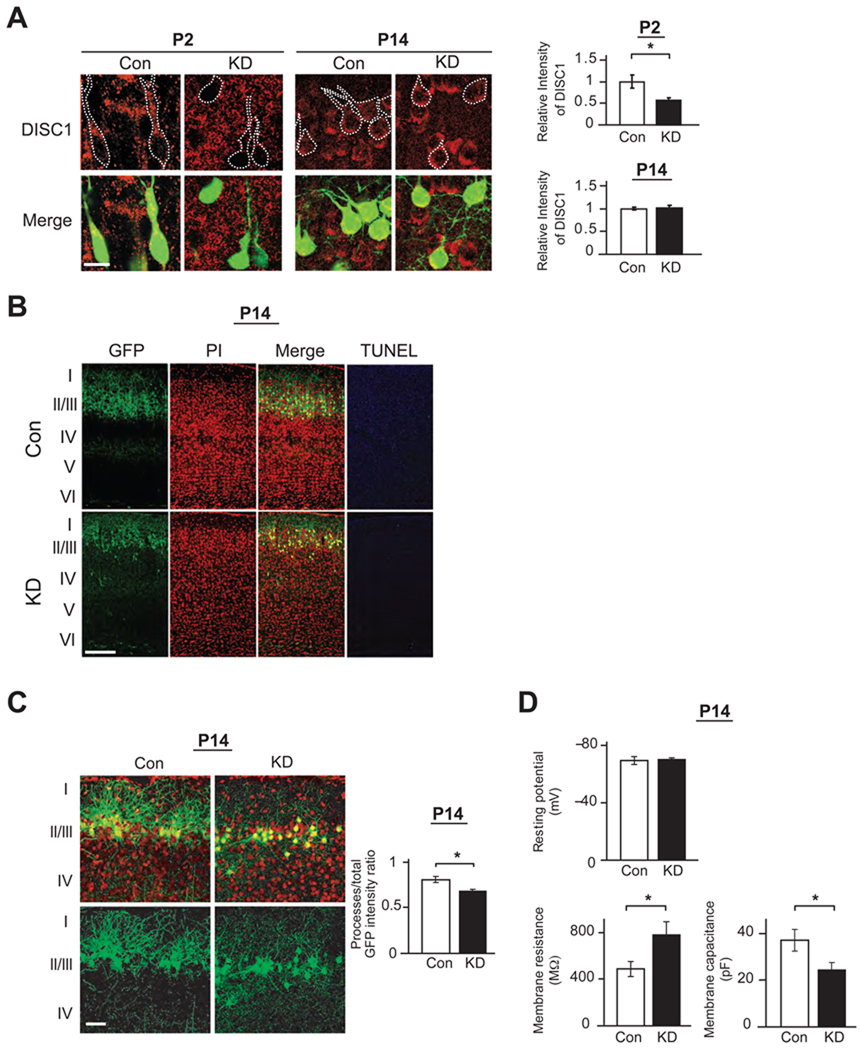
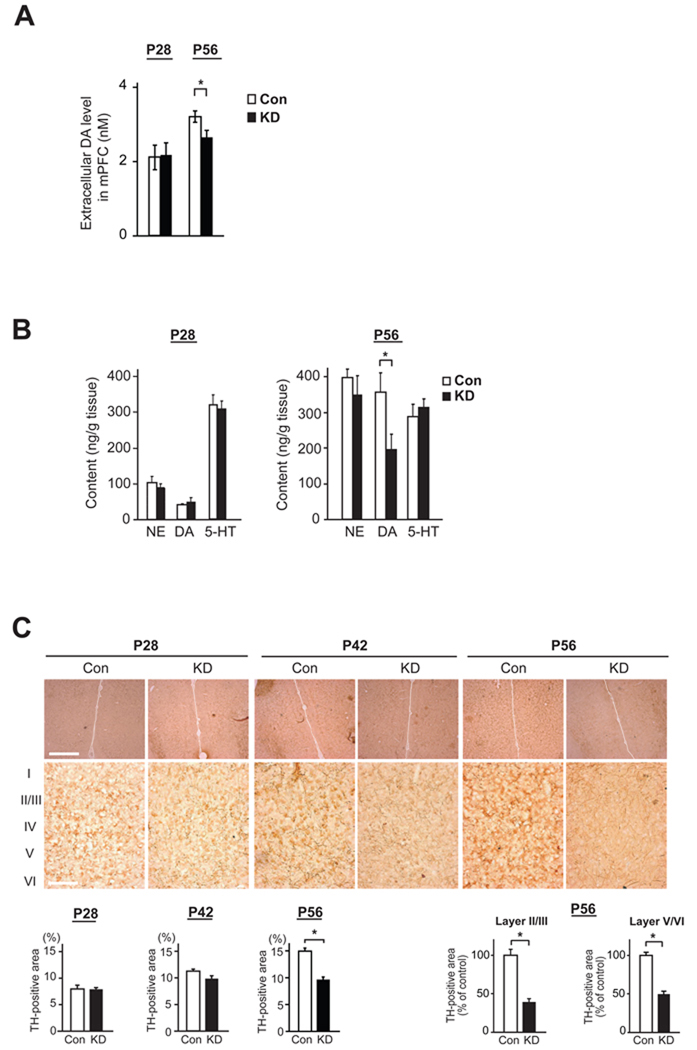
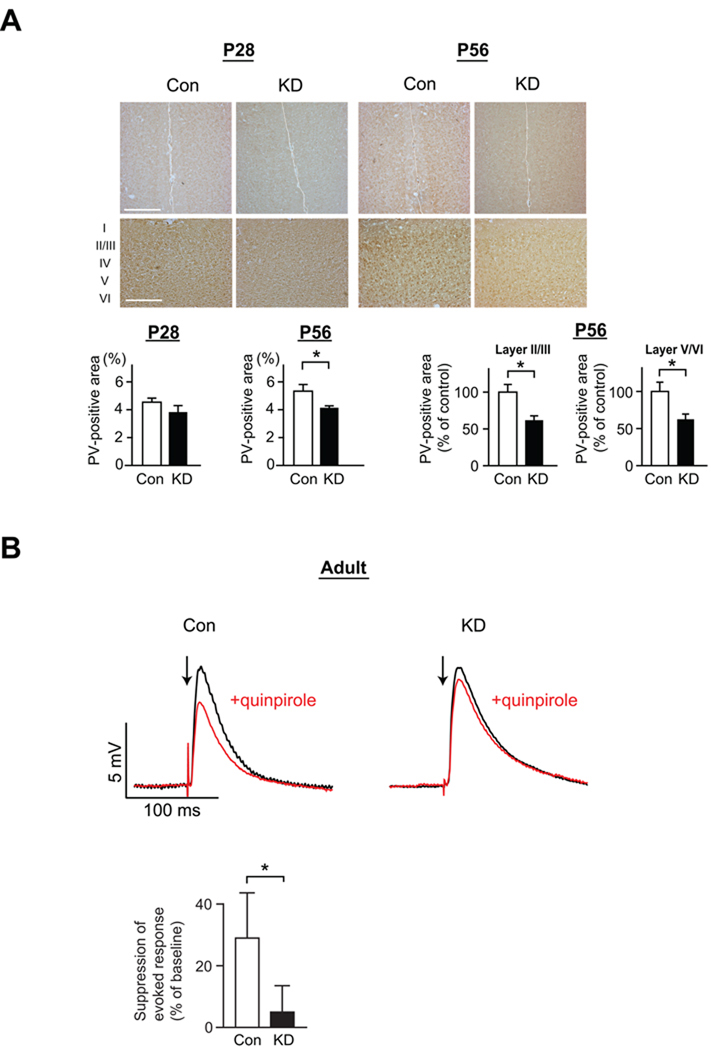
Comment in
-
Now you see it, now you don't--closing in on allostasis and developmental basis of psychiatric disorders.Neuron. 2010 Feb 25;65(4):437-9. doi: 10.1016/j.neuron.2010.02.010. Neuron. 2010. PMID: 20188647
References
-
- Akbarian S, Kim JJ, Potkin SG, Hagman JO, Tafazzoli A, Bunney WE, Jr, Jones EG. Gene expression for glutamic acid decarboxylase is reduced without loss of neurons in prefrontal cortex of schizophrenics. Arch Gen Psychiatry. 1995;52:258–266. - PubMed
-
- Akil M, Pierri JN, Whitehead RE, Edgar CL, Mohila C, Sampson AR, Lewis DA. Lamina-specific alterations in the dopamine innervation of the prefrontal cortex in schizophrenic subjects. Am J Psychiatry. 1999;156:1580–1589. - PubMed
-
- Anderson SA, Eisenstat DD, Shi L, Rubenstein JL. Interneuron migration from basal forebrain to neocortex: dependence on Dlx genes. Science. 1997;278:474–476. - PubMed
-
- Arguello PA, Gogos JA. Modeling madness in mice: one piece at a time. Neuron. 2006;52:179–196. - PubMed
-
- Benes FM, Berretta S. GABAergic interneurons: implications for understanding schizophrenia and bipolar disorder. Neuropsychopharmacology. 2001;25:1–27. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
