

Mitochondrial base excision repair assays
- PMID: 20188838
- PMCID: PMC2916069
- DOI: 10.1016/j.ymeth.2010.02.020
Mitochondrial base excision repair assays
Abstract
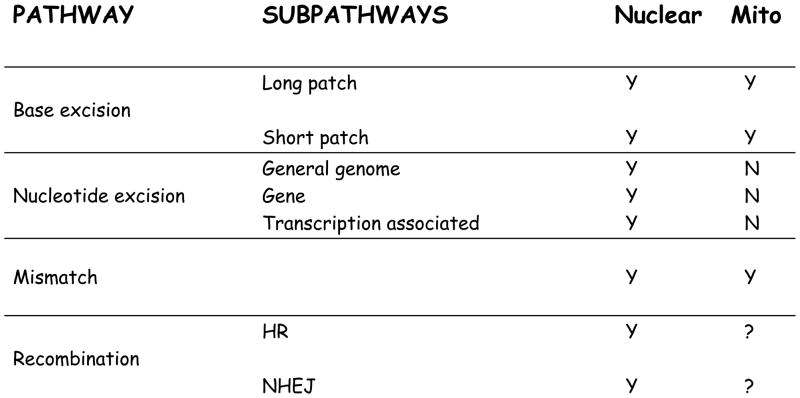
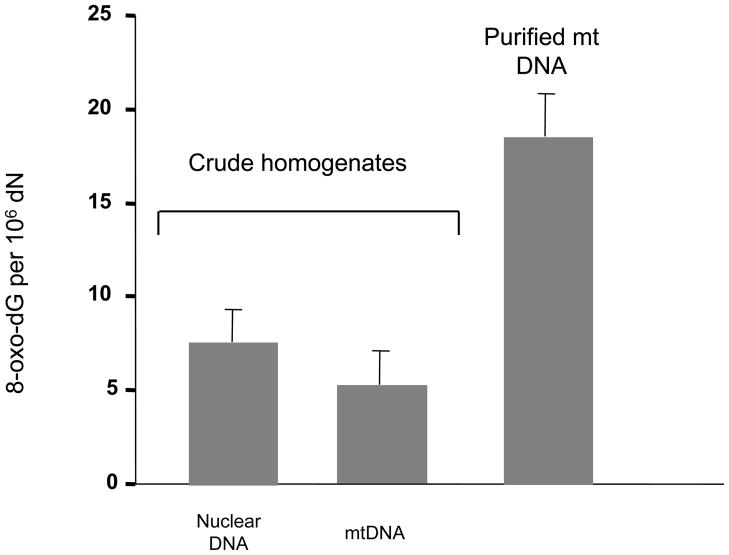
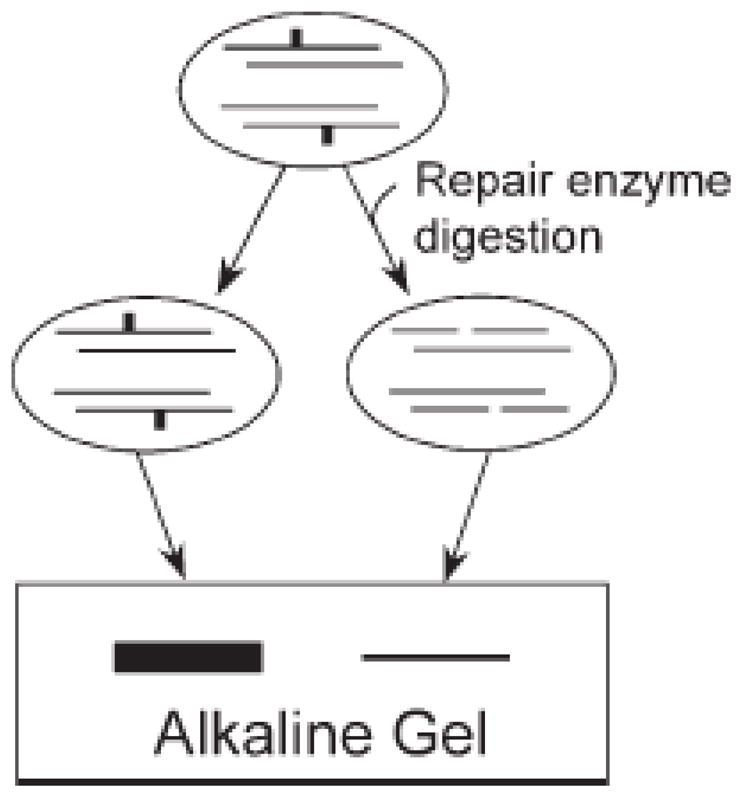
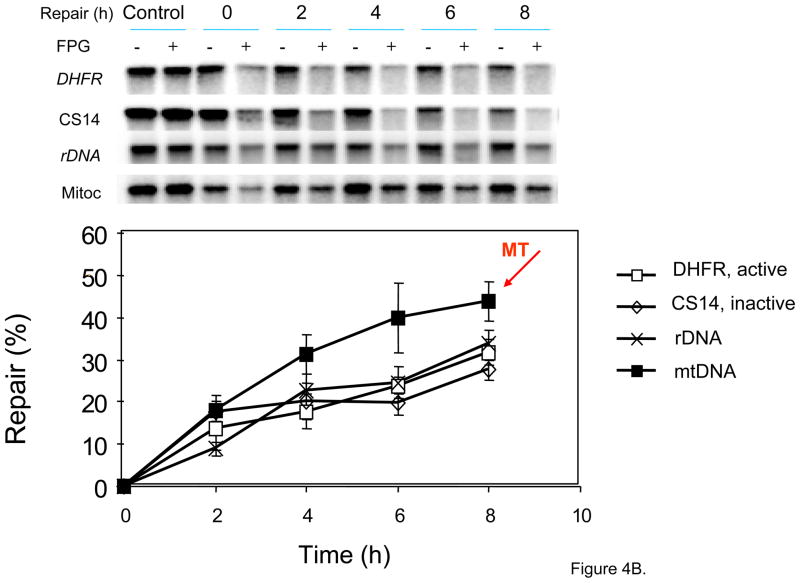
The main source of mitochondrial DNA (mtDNA) damage is reactive oxygen species (ROS) generated during normal cellular metabolism. The main mtDNA lesions generated by ROS are base modifications, such as the ubiquitous 8-oxoguanine (8-oxoG) lesion; however, base loss and strand breaks may also occur. Many human diseases are associated with mtDNA mutations and thus maintaining mtDNA integrity is critical. All of these lesions are repaired primarily by the base excision repair (BER) pathway. It is now known that mammalian mitochondria have BER, which, similarly to nuclear BER, is catalyzed by DNA glycosylases, AP endonuclease, DNA polymerase (POLgamma in mitochondria) and DNA ligase. This article outlines procedures for measuring oxidative damage formation and BER in mitochondria, including isolation of mitochondria from tissues and cells, protocols for measuring BER enzyme activities, gene-specific repair assays, chromatographic techniques as well as current optimizations for detecting 8-oxoG lesions in cells by immunofluorescence. Throughout the assay descriptions we will include methodological considerations that may help optimize the assays in terms of resolution and repeatability.
Published by Elsevier Inc.
Figures

Similar articles
-
Expression changes in DNA repair enzymes and mitochondrial DNA damage in aging rat lens.Mol Vis. 2010 Aug 27;16:1754-63. Mol Vis. 2010. PMID: 20808729 Free PMC article.
-
DNA base excision repair activities and pathway function in mitochondrial and cellular lysates from cells lacking mitochondrial DNA.Nucleic Acids Res. 2004 Apr 23;32(7):2181-92. doi: 10.1093/nar/gkh533. Print 2004. Nucleic Acids Res. 2004. PMID: 15107486 Free PMC article.
-
In vitro measurement of DNA base excision repair in isolated mitochondria.Methods Mol Biol. 2009;554:213-31. doi: 10.1007/978-1-59745-521-3_14. Methods Mol Biol. 2009. PMID: 19513677
-
Early steps in the DNA base excision/single-strand interruption repair pathway in mammalian cells.Cell Res. 2008 Jan;18(1):27-47. doi: 10.1038/cr.2008.8. Cell Res. 2008. PMID: 18166975 Free PMC article. Review.
-
Mitochondrial repair of 8-oxoguanine and changes with aging.Exp Gerontol. 2002 Oct-Nov;37(10-11):1189-96. doi: 10.1016/s0531-5565(02)00142-0. Exp Gerontol. 2002. PMID: 12470830 Review.
Cited by
-
Arginine methylation of APE1 promotes its mitochondrial translocation to protect cells from oxidative damage.Free Radic Biol Med. 2020 Oct;158:60-73. doi: 10.1016/j.freeradbiomed.2020.06.027. Epub 2020 Jul 15. Free Radic Biol Med. 2020. PMID: 32679368 Free PMC article.
-
Aprataxin localizes to mitochondria and preserves mitochondrial function.Proc Natl Acad Sci U S A. 2011 May 3;108(18):7437-42. doi: 10.1073/pnas.1100084108. Epub 2011 Apr 18. Proc Natl Acad Sci U S A. 2011. PMID: 21502511 Free PMC article.
-
Interaction between RECQL4 and OGG1 promotes repair of oxidative base lesion 8-oxoG and is regulated by SIRT1 deacetylase.Nucleic Acids Res. 2020 Jul 9;48(12):6530-6546. doi: 10.1093/nar/gkaa392. Nucleic Acids Res. 2020. PMID: 32432680 Free PMC article.
-
Slow mitochondrial repair of 5'-AMP renders mtDNA susceptible to damage in APTX deficient cells.Sci Rep. 2015 Aug 10;5:12876. doi: 10.1038/srep12876. Sci Rep. 2015. PMID: 26256098 Free PMC article.
-
Maternal transmission of mitochondrial diseases.Genet Mol Biol. 2020 Mar 2;43(1 suppl. 1):e20190095. doi: 10.1590/1678-4685-GMB-2019-0095. eCollection 2020. Genet Mol Biol. 2020. PMID: 32141474 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
