

Retinoids regulate a developmental checkpoint for tissue regeneration in Drosophila
- PMID: 20189388
- PMCID: PMC2847081
- DOI: 10.1016/j.cub.2010.01.038
Retinoids regulate a developmental checkpoint for tissue regeneration in Drosophila
Abstract
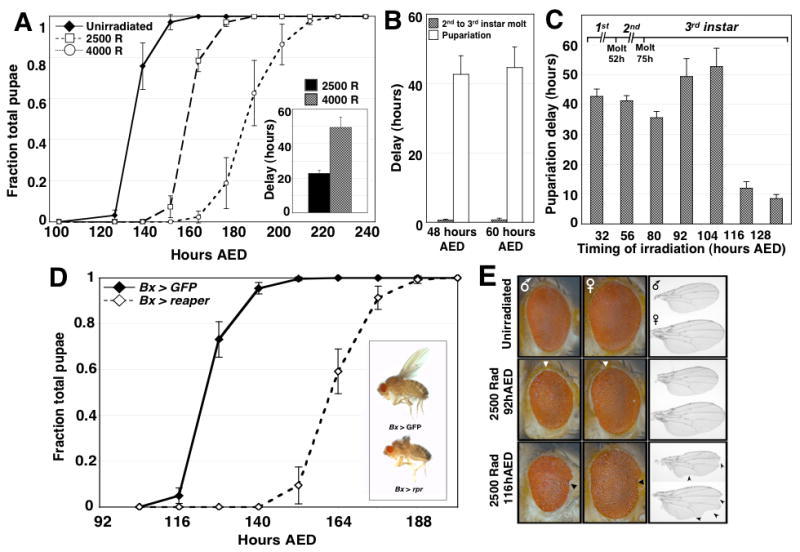
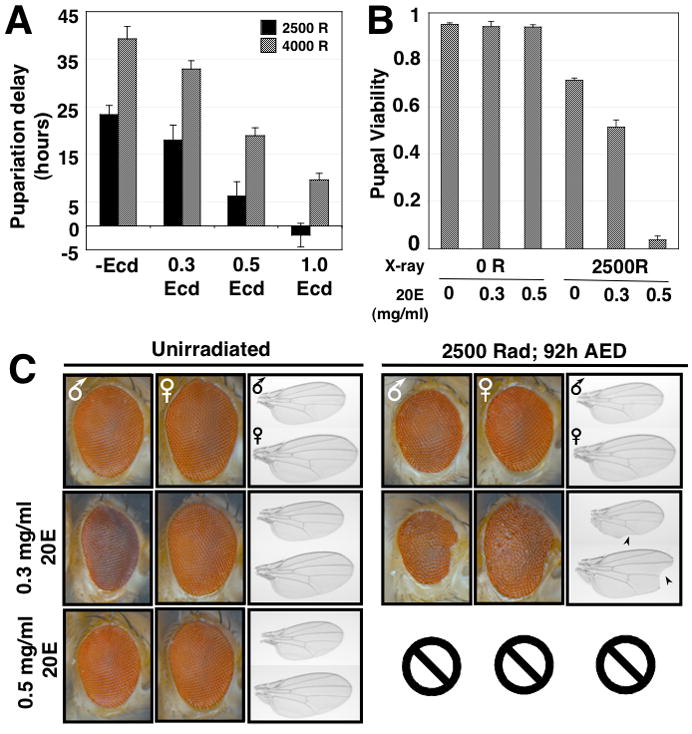
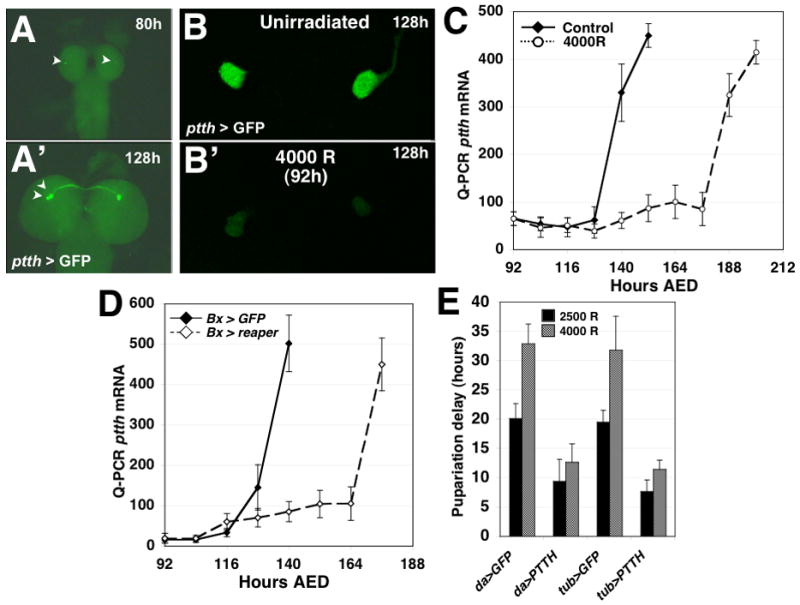
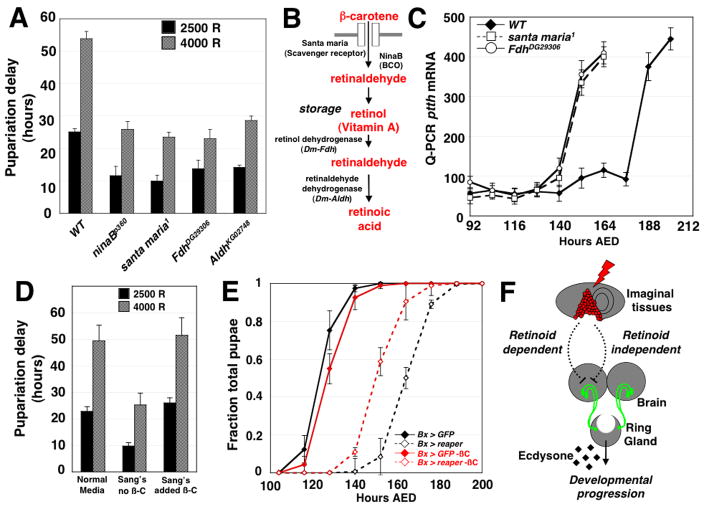
Damage to Drosophila imaginal discs elicits a robust regenerative response from the surviving tissue [1-4]. However, as in other organisms, developmental progression and differentiation can restrict the regenerative capacity of Drosophila tissues. Experiments in Drosophila and other holometabolous insects have demonstrated that either damage to imaginal tissues [5, 6] or transplantation of a damaged imaginal disc [7, 8] delays the onset of metamorphosis. Therefore, in Drosophila there appears to be a mechanism that senses tissue damage and extends the larval phase to coordinate tissue regeneration with the overall developmental program of the organism. However, how such a pathway functions remains unknown. Here we demonstrate that a developmental checkpoint extends larval growth after imaginal disc damage by inhibiting the transcription of the gene encoding PTTH, a neuropeptide that promotes the release of the steroid hormone ecdysone. Using a genetic screen, we identify a previously unsuspected role for retinoid biosynthesis in regulating PTTH expression and delaying development in response to tissue damage. Retinoid signaling plays an important but poorly defined role in several vertebrate regeneration models [9-11]. Our findings demonstrate that retinoid biosynthesis in Drosophila is important for the maintenance of a condition that is permissive for regenerative growth.
Figures
References
-
- Bryant P. Regeneration and duplication in imaginal discs. Ciba Foundation symposium.1975. - PubMed
-
- Bryant P. Pattern formation in the imaginal wing disc of Drosophila melanogaster: fate map, regeneration and duplication. J Exp Zool. 1975;193:49–77. - PubMed
-
- Schubiger G. Regeneration, duplication and transdetermination in fragments of the leg disc of Drosophila melanogaster. Dev Biol. 1971;26:277–295. - PubMed
-
- Hussey R, Thompson W, Calhoun E. The influence of x-rays on the development of drosophila larvae. Science. 1927;66:65–66. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
