

Functional diversity of four glycoside hydrolase family 3 enzymes from the rumen bacterium Prevotella bryantii B14
- PMID: 20190048
- PMCID: PMC2863481
- DOI: 10.1128/JB.01654-09
Functional diversity of four glycoside hydrolase family 3 enzymes from the rumen bacterium Prevotella bryantii B14
Abstract
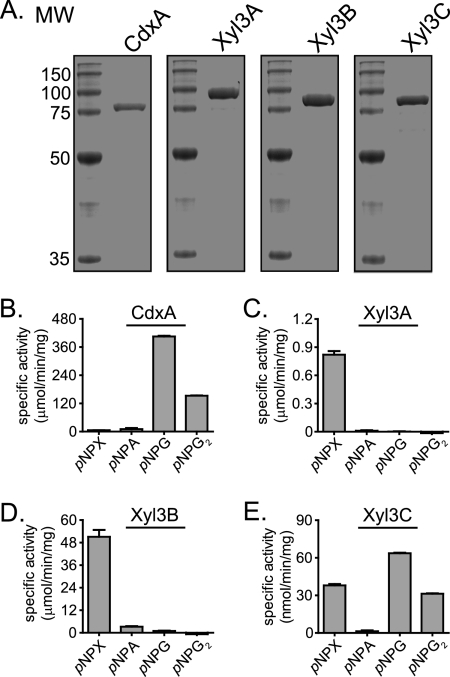
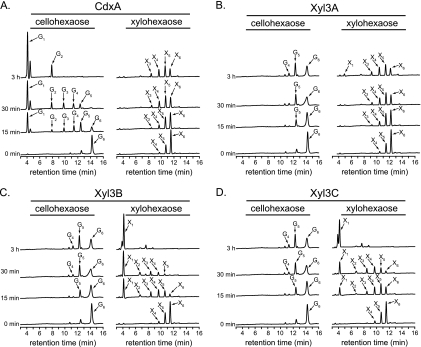
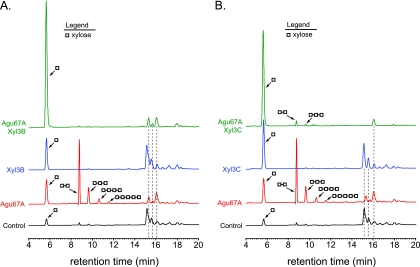
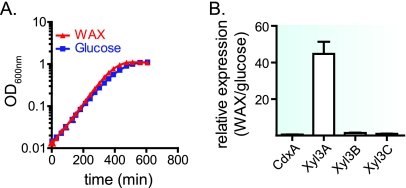
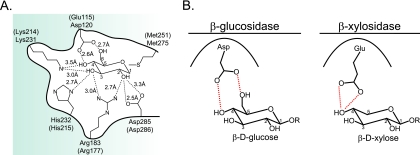
Prevotella bryantii B(1)4 is a member of the phylum Bacteroidetes and contributes to the degradation of hemicellulose in the rumen. The genome of P. bryantii harbors four genes predicted to encode glycoside hydrolase (GH) family 3 (GH3) enzymes. To evaluate whether these genes encode enzymes with redundant biological functions, each gene was cloned and expressed in Escherichia coli. Biochemical analysis of the recombinant proteins revealed that the enzymes exhibit different substrate specificities. One gene encoded a cellodextrinase (CdxA), and three genes encoded beta-xylosidase enzymes (Xyl3A, Xyl3B, and Xyl3C) with different specificities for either para-nitrophenyl (pNP)-linked substrates or substituted xylooligosaccharides. To identify the amino acid residues that contribute to catalysis and substrate specificity within this family of enzymes, the roles of conserved residues (R177, K214, H215, M251, and D286) in Xyl3B were probed by site-directed mutagenesis. Each mutation led to a severely decreased catalytic efficiency without a change in the overall structure of the mutant enzymes. Through amino acid sequence alignments, an amino acid residue (E115) that, when mutated to aspartic acid, resulted in a 14-fold decrease in the k(cat)/K(m) for pNP-beta-d-xylopyranoside (pNPX) with a concurrent 1.1-fold increase in the k(cat)/K(m) for pNP-beta-d-glucopyranoside (pNPG) was identified. Amino acid residue E115 may therefore contribute to the discrimination between beta-xylosides and beta-glucosides. Our results demonstrate that each of the four GH3 enzymes has evolved to perform a specific role in lignopolysaccharide hydrolysis and provide insight into the role of active-site residues in catalysis and substrate specificity for GH3 enzymes.
Figures
References
-
- Aziz, R. K., D. Bartels, A. A. Best, M. DeJongh, T. Disz, R. A. Edwards, K. Formsma, S. Gerdes, E. M. Glass, M. Kubal, F. Meyer, G. J. Olsen, R. Olson, A. L. Osterman, R. A. Overbeek, L. K. McNeil, D. Paarmann, T. Paczian, B. Parrello, G. D. Pusch, C. Reich, R. Stevens, O. Vassieva, V. Vonstein, A. Wilke, and O. Zagnitko. 2008. The RAST server: rapid annotations using subsystems technology. BMC Genomics 9:75. - PMC - PubMed
-
- Bause, E., and G. Legler. 1974. Isolation and amino acid sequence of a hexadecapeptide from the active site of beta-glucosidase A3 from Aspergillus wentii. Hoppe-Seylers Z. Physiol. Chem. 355:438-442. - PubMed
-
- Brulc, J. M., D. A. Antonopoulos, M. E. Miller, M. K. Wilson, A. C. Yannarell, E. A. Dinsdale, R. E. Edwards, E. D. Frank, J. B. Emerson, P. Wacklin, P. M. Coutinho, B. Henrissat, K. E. Nelson, and B. A. White. 2009. Gene-centric metagenomics of the fiber-adherent bovine rumen microbiome reveals forage specific glycoside hydrolases. Proc. Natl. Acad. Sci. U. S. A. 106:1948-1953. - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
