

Release of adenosine and ATP during ischemia and epilepsy
- PMID: 20190959
- PMCID: PMC2769001
- DOI: 10.2174/157015909789152146
Release of adenosine and ATP during ischemia and epilepsy
Abstract
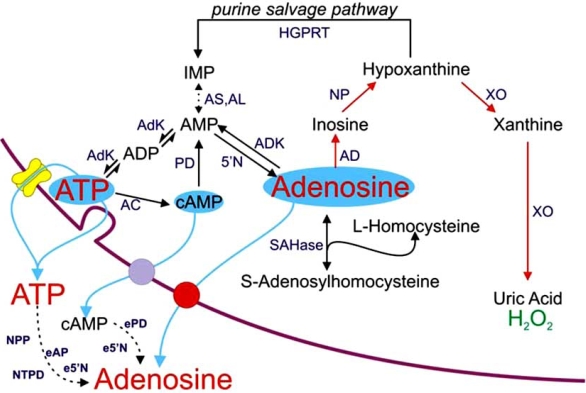
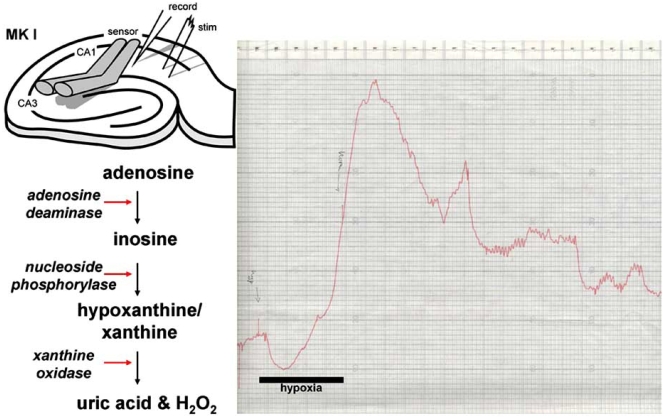
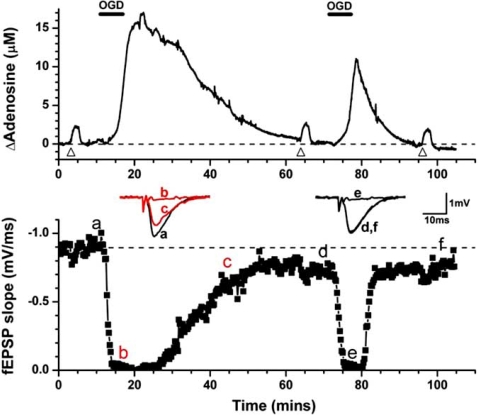
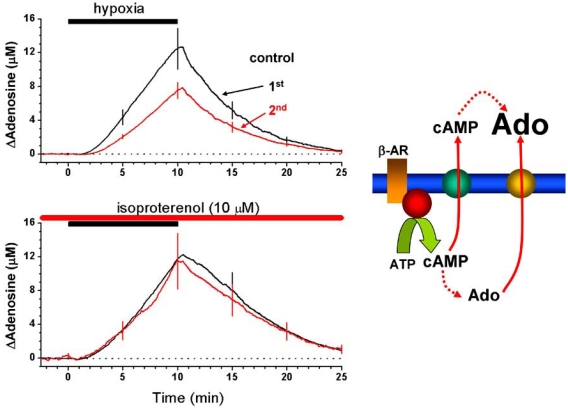
Eighty years ago Drury & Szent-Györgyi described the actions of adenosine, AMP (adenylic acid) and ATP (pyrophosphoric or diphosphoric ester of adenylic acid) on the mammalian cardiovascular system, skeletal muscle, intestinal and urinary systems. Since then considerable insight has been gleaned on the means by which these compounds act, not least of which in the distinction between the two broad classes of their respective receptors, with their many subtypes, and the ensuing diversity in cellular consequences their activation invokes. These myriad actions are of course predicated on the release of the purines into the extracellular milieu, but, surprisingly, there is still considerable ambiguity as to how this occurs in various physiological and pathophysiological conditions. In this review we summarise the release of ATP and adenosine during seizures and cerebral ischemia and discuss mechanisms by which the purines adenosine and ATP may be released from cells in the CNS under these conditions.
Figures
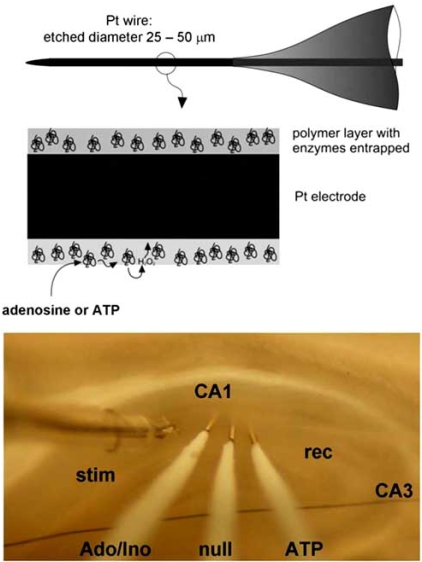
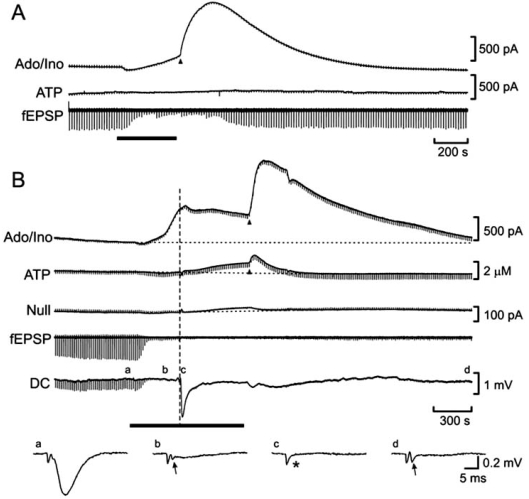
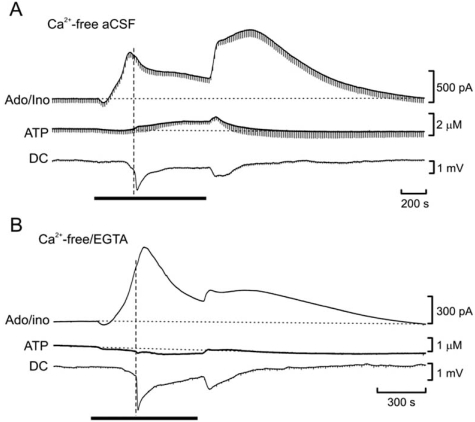

References
-
- Abbracchio MP, Boeynaems JM, Barnard EA, Boyer JL, Kennedy C, Miras-Portugal MT, King BF, Gachet C, Jacobson KA, Weisman GA, Burnstock G. Characterization of the UDP-glucose receptor (re-named here the P2Y14 receptor) adds diversity to the P2Y receptor family. Trends Pharmacol. Sci. 2003;24:52–55. - PMC - PubMed
-
- Abbracchio MP, Burnstock G, Boeynaems JM, Barnard EA, Boyer JL, Kennedy C, Knight GE, Fumagalli M, Gachet C, Jacobson KA, Weisman GA. International Union of Pharmacology LVIII: update on the P2Y G protein-coupled nucleotide receptors: from molecular mechanisms and pathophysiology to therapy. Pharmacol. Rev. 2006;58:281–341. - PMC - PubMed
-
- Abbracchio MP, Burnstock G, Verkhratsky A, Zimmermann H. Purinergic signalling in the nervous system: an overview. Trends Neurosci. 2009;32:19–29. - PubMed
-
- Bao L, Locovei S, Dahl G. Pannexin membrane channels are mechanosensitive conduits for ATP. FEBS Lett. 2004;572:65–68. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources