

Lessons learned from UvrD helicase: mechanism for directional movement
- PMID: 20192763
- PMCID: PMC3480338
- DOI: 10.1146/annurev.biophys.093008.131415
Lessons learned from UvrD helicase: mechanism for directional movement
Abstract
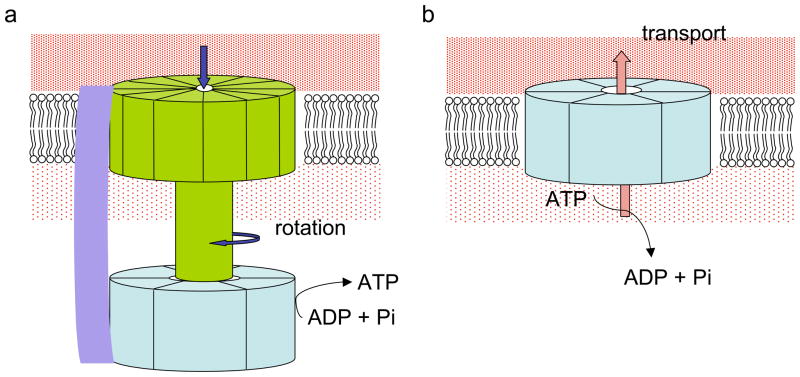
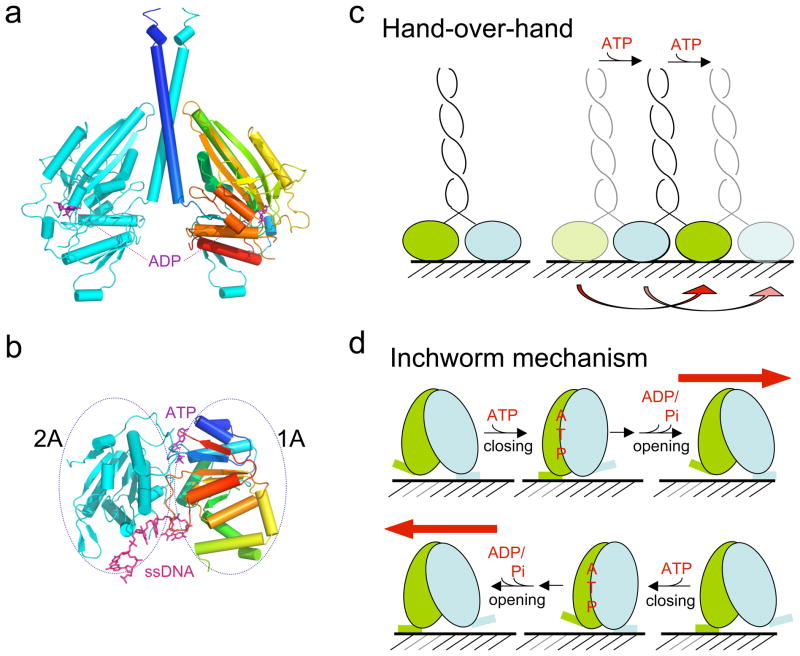
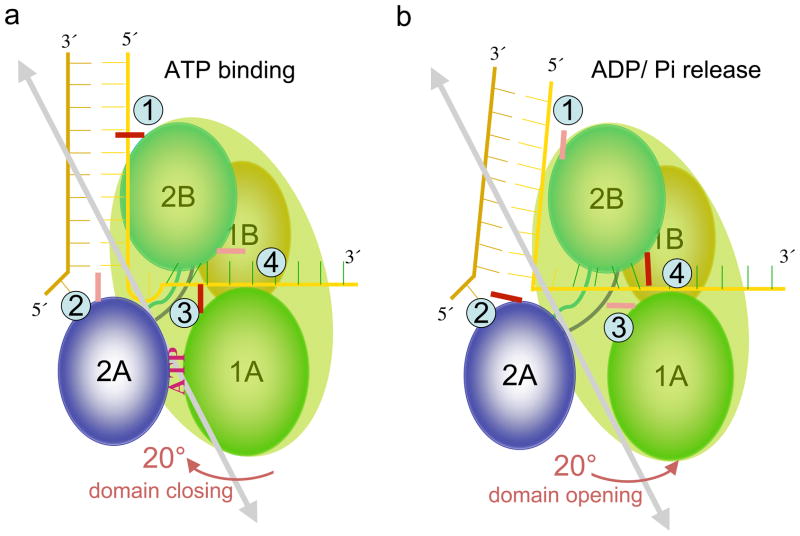
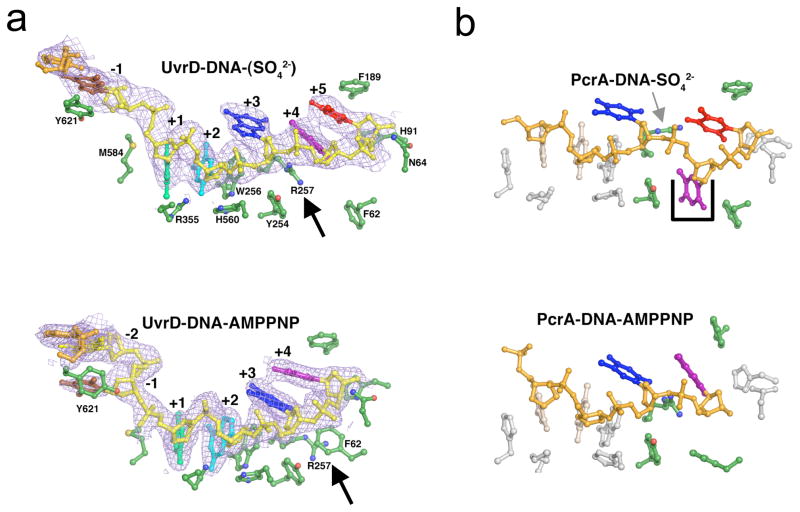
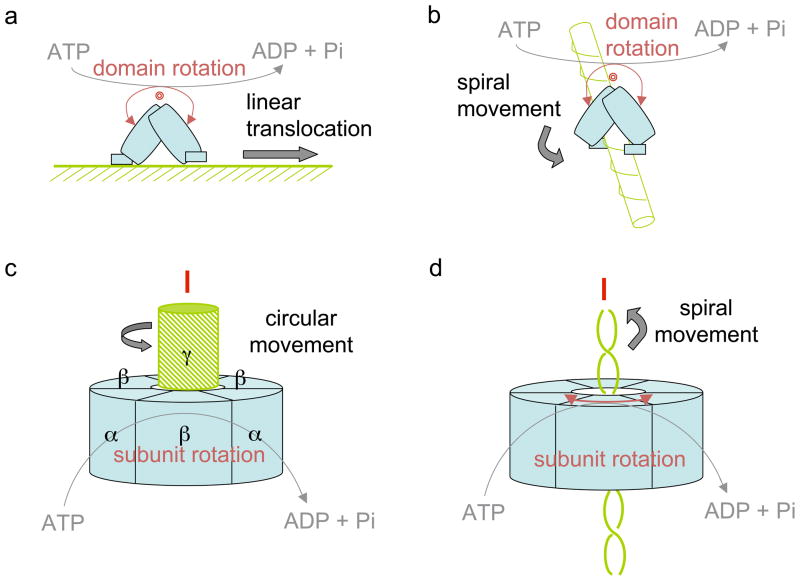
How do molecular motors convert chemical energy to mechanical work? Helicases and nucleic acids offer simple motor systems for extensive biochemical and biophysical analyses. Atomic resolution structures of UvrD-like helicases complexed with DNA in the presence of AMPPNP, ADP.Pi, and Pi reveal several salient points that aid our understanding of mechanochemical coupling. Each ATPase cycle causes two motor domains to rotationally close and open. At a minimum, two motor-track contact points of alternating tight and loose attachment convert domain rotations to unidirectional movement. A motor is poised for action only when fully in contact with its track and, if applicable, working against a load. The orientation of domain rotation relative to the track determines whether the movement is linear, spiral, or circular. Motors powered by ATPases likely deliver each power stroke in two parts, before and after ATP hydrolysis. Implications of these findings for analyzing hexameric helicase, F(1)F(0) ATPase, and kinesin are discussed.
Figures
Similar articles
-
Rotations of the 2B sub-domain of E. coli UvrD helicase/translocase coupled to nucleotide and DNA binding.J Mol Biol. 2011 Aug 19;411(3):633-48. doi: 10.1016/j.jmb.2011.06.019. Epub 2011 Jun 17. J Mol Biol. 2011. PMID: 21704638 Free PMC article.
-
Study of the ATP-binding site of helicase IV from Escherichia coli.Biochem Biophys Res Commun. 2006 Mar 17;341(3):828-36. doi: 10.1016/j.bbrc.2006.01.040. Epub 2006 Jan 23. Biochem Biophys Res Commun. 2006. PMID: 16442499
-
A two-site mechanism for ATP hydrolysis by the asymmetric Rep dimer P2S as revealed by site-specific inhibition with ADP-A1F4.Biochemistry. 1997 Mar 18;36(11):3115-25. doi: 10.1021/bi9621977. Biochemistry. 1997. PMID: 9115987
-
On helicases and other motor proteins.Curr Opin Struct Biol. 2008 Apr;18(2):243-57. doi: 10.1016/j.sbi.2008.01.007. Epub 2008 Mar 10. Curr Opin Struct Biol. 2008. PMID: 18329872 Free PMC article. Review.
-
Roles of the C-Terminal Amino Acids of Non-Hexameric Helicases: Insights from Escherichia coli UvrD.Int J Mol Sci. 2021 Jan 20;22(3):1018. doi: 10.3390/ijms22031018. Int J Mol Sci. 2021. PMID: 33498436 Free PMC article. Review.
Cited by
-
Structural and mechanistic insight into DNA unwinding by Deinococcus radiodurans UvrD.PLoS One. 2013 Oct 15;8(10):e77364. doi: 10.1371/journal.pone.0077364. eCollection 2013. PLoS One. 2013. PMID: 24143224 Free PMC article.
-
The conserved C-terminus of the PcrA/UvrD helicase interacts directly with RNA polymerase.PLoS One. 2013 Oct 16;8(10):e78141. doi: 10.1371/journal.pone.0078141. eCollection 2013. PLoS One. 2013. PMID: 24147116 Free PMC article.
-
Investigation of bacterial nucleotide excision repair using single-molecule techniques.DNA Repair (Amst). 2014 Aug;20:41-48. doi: 10.1016/j.dnarep.2013.10.012. Epub 2014 Jan 25. DNA Repair (Amst). 2014. PMID: 24472181 Free PMC article. Review.
-
DNA Mismatch Repair.EcoSal Plus. 2012 Nov;5(1):10.1128/ecosalplus.7.2.5. doi: 10.1128/ecosalplus.7.2.5. EcoSal Plus. 2012. PMID: 26442827 Free PMC article.
-
The Role of SF1 and SF2 Helicases in Biotechnological Applications.Appl Biochem Biotechnol. 2024 Dec;196(12):9064-9084. doi: 10.1007/s12010-024-05027-w. Epub 2024 Aug 2. Appl Biochem Biotechnol. 2024. PMID: 39093351 Review.
References
-
- Abrahams JP, Leslie AG, Lutter R, Walker JE. Structure at 2.8 A resolution of F1-ATPase from bovine heart mitochondria. Nature. 1994;370:621–8. - PubMed
-
- Ali JA, Lohman TM. Kinetic measurement of the step size of DNA unwinding by Escherichia coli UvrD helicase. Science. 1997;275:377–80. - PubMed
-
- Amos LA, van den Ent F, Lowe J. Structural/functional homology between the bacterial and eukaryotic cytoskeletons. Curr Opin Cell Biol. 2004;16:24–31. - PubMed
-
- Andersen CB, Ballut L, Johansen JS, Chamieh H, Nielsen KH, et al. Structure of the exon junction core complex with a trapped DEAD-box ATPase bound to RNA. Science. 2006;313:1968–72. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
