

Eukaryotic chemotaxis: a network of signaling pathways controls motility, directional sensing, and polarity
- PMID: 20192768
- PMCID: PMC4364543
- DOI: 10.1146/annurev.biophys.093008.131228
Eukaryotic chemotaxis: a network of signaling pathways controls motility, directional sensing, and polarity
Abstract
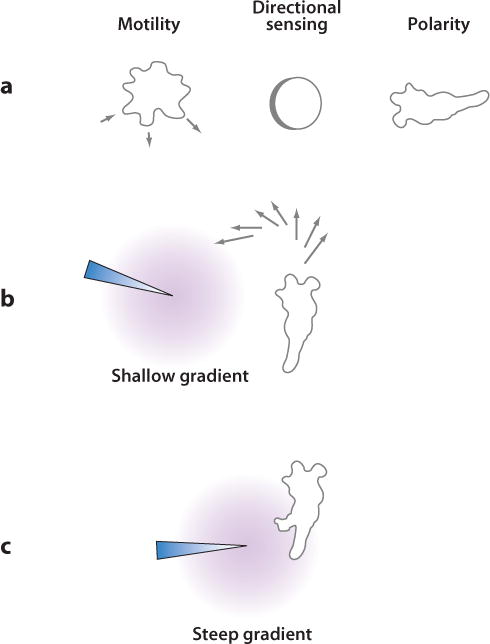
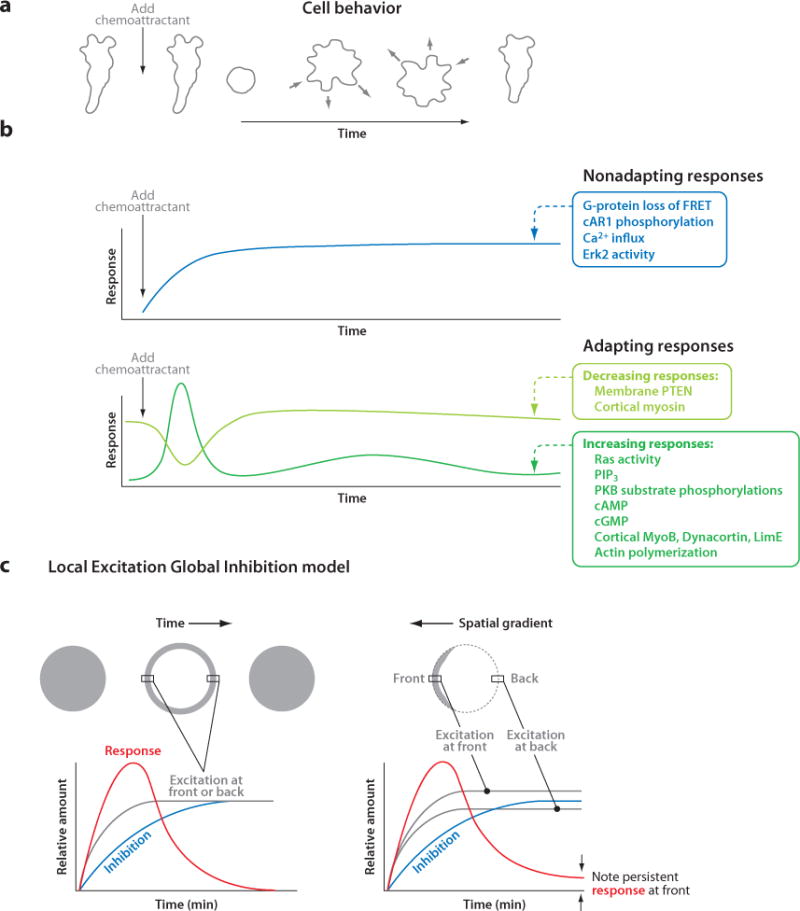
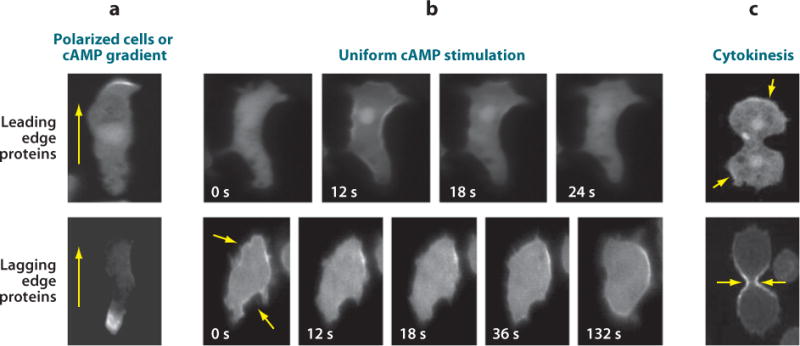
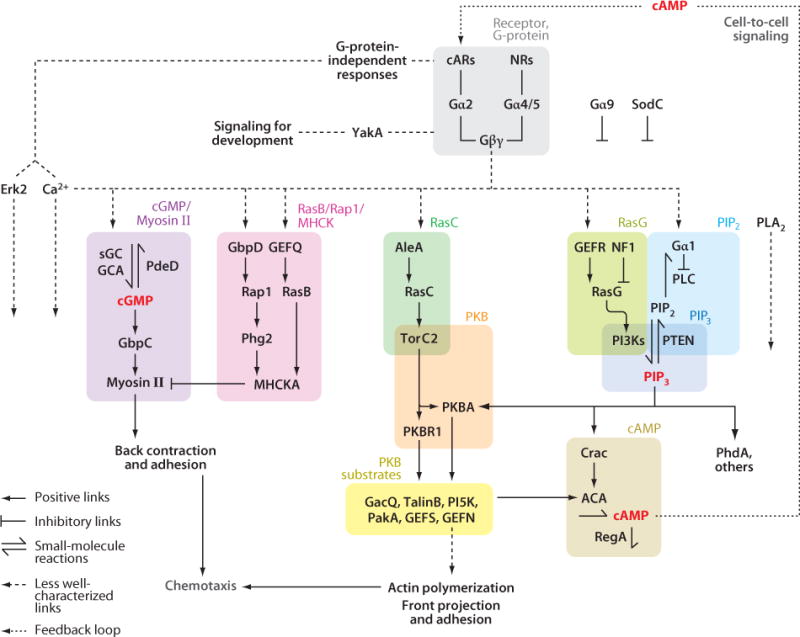
Chemotaxis, the directed migration of cells in chemical gradients, is a vital process in normal physiology and in the pathogenesis of many diseases. Chemotactic cells display motility, directional sensing, and polarity. Motility refers to the random extension of pseudopodia, which may be driven by spontaneous actin waves that propagate through the cytoskeleton. Directional sensing is mediated by a system that detects temporal and spatial stimuli and biases motility toward the gradient. Polarity gives cells morphologically and functionally distinct leading and lagging edges by relocating proteins or their activities selectively to the poles. By exploiting the genetic advantages of Dictyostelium, investigators are working out the complex network of interactions between the proteins that have been implicated in the chemotactic processes of motility, directional sensing, and polarity.
Figures
References
LITERATURE CITED
-
- Andrew N, Insall RH. Chemotaxis in shallow gradients is mediated independently of PtdIns 3-kinase by biased choices between random protrusions. Nat Cell Biol. 2007;9:193–200. - PubMed
-
- Blaser H, Reichman-Fried M, Castanon I, Dumstrei K, Marlow FL, et al. Migration of zebrafish primordial germ cells: a role for myosin contraction and cytoplasmic flow. Dev Cell. 2006;11:613–27. - PubMed
-
- Bolourani P, Spiegelman GB, Weeks G. Rap1 activation in response to cAMP occurs downstream of Ras activation during Dictyostelium aggregation. J Biol Chem. 2008;283:10232–40. - PubMed
RELATED RESOURCES
-
- Franca-Koh J, Kamimura Y, Devreotes P. Navigating signaling networks: chemotaxis in Dictyostelium discoideum. Curr Opin Genet Dev. 2006;16:333–38. - PubMed
-
- Kay RR, Langridge P, Traynor D, Hoeller O. Changing directions in the study of chemotaxis. Nat Rev Mol Cell Biol. 2008;9:455–63. - PubMed
-
- Van Haastert PJ, Veltman DM. Chemotaxis: navigating by multiple signaling pathways. Sci STKE. 2007:pe40. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
