

Maximum yields of microsomal-type membranes from small amounts of plant material without requiring ultracentrifugation
- PMID: 20193653
- PMCID: PMC3685806
- DOI: 10.1016/j.ab.2010.02.030
Maximum yields of microsomal-type membranes from small amounts of plant material without requiring ultracentrifugation
Abstract
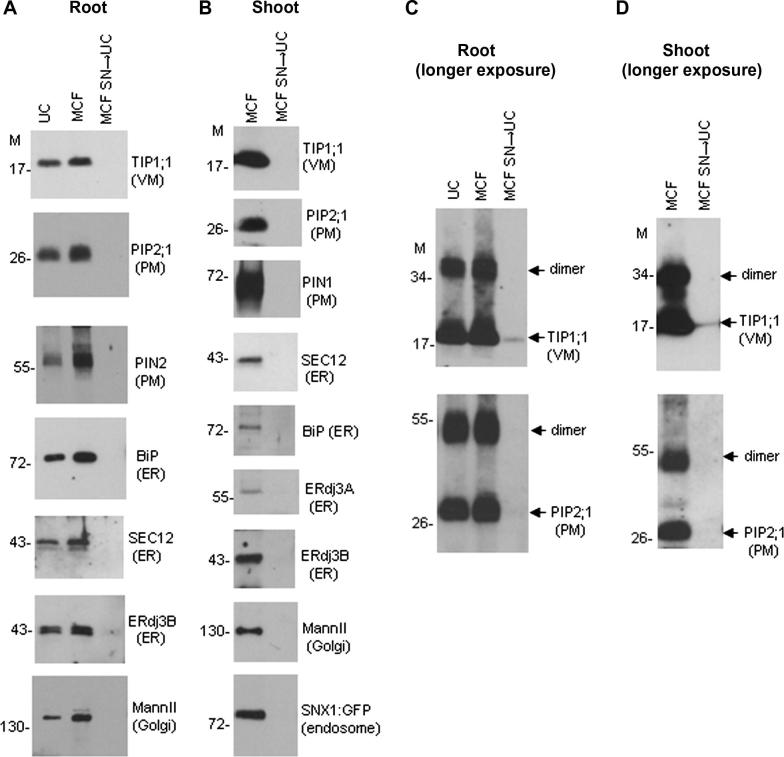
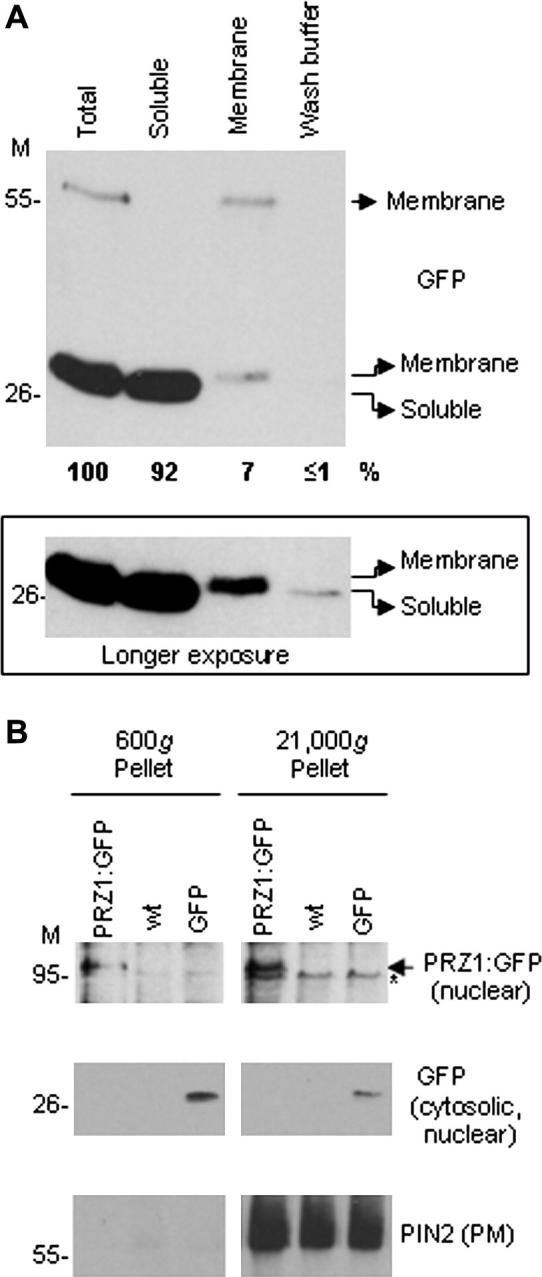
Isolation of a microsomal membrane fraction is a common procedure in studies involving membrane proteins. By conventional definition, microsomal membranes are collected by centrifugation of a postmitochondrial fraction at 100,000g in an ultracentrifuge, a method originally developed for large amounts of mammalian tissue. We present a method for isolating microsomal-type membranes from small amounts of Arabidopsis thaliana plant material that does not rely on ultracentrifugation but instead uses the lower relative centrifugal force (21,000g) of a microcentrifuge. We show that the 21,000g pellet is equivalent to that obtained at 100,000g and that it contains all of the membrane fractions expected in a conventional microsomal fraction. Our method incorporates specific manipulation of sample density throughout the procedure, with minimal preclearance, minimal volumes of extraction buffer, and minimal sedimentation pathlength. These features allow maximal membrane yields, enabling membrane isolation from limited amounts of material. We further demonstrate that conventional ultracentrifuge-based protocols give submaximal yields due to losses during early stages of the procedure; that is, extensive amounts of microsomal-type membranes can sediment prematurely during the typical preclearance steps. Our protocol avoids such losses, thereby ensuring maximal yield and a representative total membrane fraction. The principles of our method can be adapted for nonplant material.
Copyright (c) 2010 Elsevier Inc. All rights reserved.
Figures





References
-
- Reid E., Williamson R. Centrifugation. Methods Enzymol. 1974;31:713–733. - PubMed
-
- Graham J. Isolation of subcellular organelles and membranes. In: Rickwood D., editor. Centrifugation: A Practical Approach. second ed. IRL Press; Oxford, UK: 1984. pp. 161–182.
-
- Ozols J. Preparation of membrane fractions. Methods Enzymol. 1990;182:225–235. - PubMed
-
- Robinson D.G., Hinz G., Oberbeck K. Isolation of endo- and plasma membranes. In: Harris N., Oparka K.J., editors. Plant Cell Biology: A Practical Approach. Oxford University Press; Oxford, UK: 1994. pp. 245–272.
-
- Dey P.M., Brownleader M.D., Harborne J.B. The plant, the cell, and its molecular components. In: Dey P.M., Harborne J.B., editors. Plant Biochemistry. Academic Press; San Diego: 1997. pp. 1–47.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
