

Hedgehog signaling induces arterial endothelial cell formation by repressing venous cell fate
- PMID: 20193674
- PMCID: PMC3197743
- DOI: 10.1016/j.ydbio.2010.02.028
Hedgehog signaling induces arterial endothelial cell formation by repressing venous cell fate
Abstract
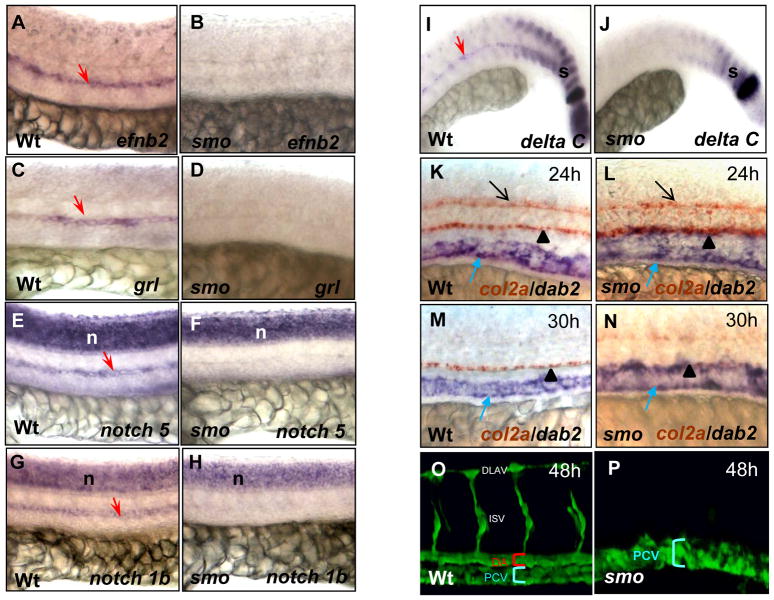
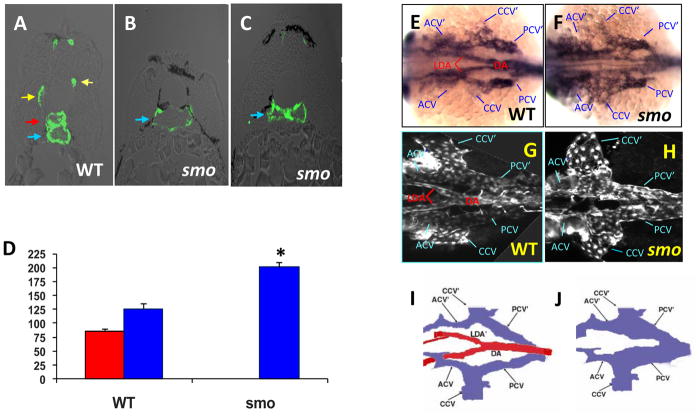
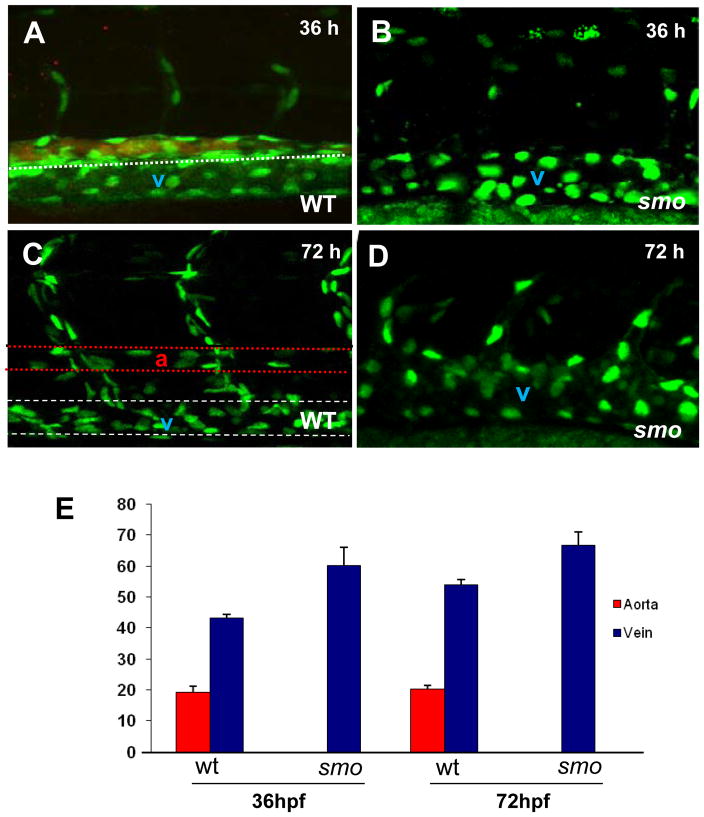
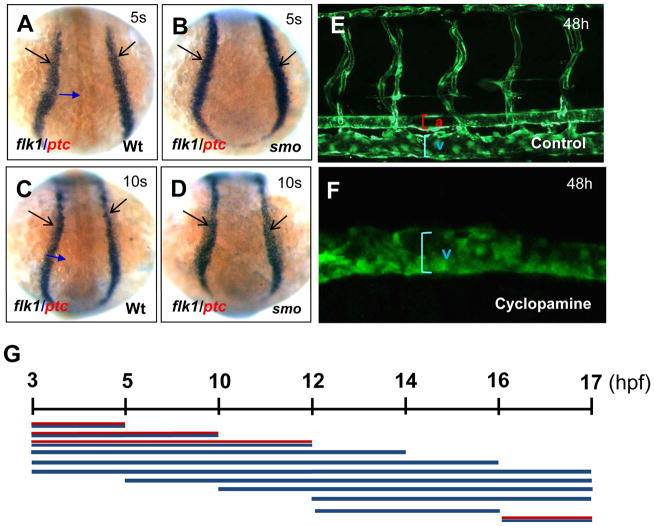
In vertebrate embryos, the dorsal aorta and the posterior cardinal vein form in the trunk to comprise the original circulatory loop. Previous studies implicate Hedgehog (Hh) signaling in the development of the dorsal aorta. However, the mechanism controlling specification of artery versus vein remains unclear. Here, we investigated the cell-autonomous mechanism of Hh signaling in angioblasts (endothelial progenitor cells) during arterial-venous specification utilizing zebrafish mutations in Smoothened (Smo), a G protein-coupled receptor essential for Hh signaling. smo mutants exhibit an absence of the dorsal aorta accompanied by a reciprocal expansion of the posterior cardinal vein. The increased number of venous cells is equivalent to the loss of arterial cells in embryos with loss of Smo function. Activation of Hh signaling expands the arterial cell population at the expense of venous cell fate. Time-lapse imaging reveals two sequential waves of migrating progenitor cells that contribute to the dorsal aorta and the posterior cardinal vein, respectively. Angioblasts deficient in Hh signaling fail to contribute to the arterial wave; instead, they all migrate medially as a single population to form the venous wave. Cell transplantation analyses demonstrate that Smo plays a cell-autonomous role in specifying angioblasts to become arterial cells, and Hh signaling-depleted angioblasts differentiate into venous cells instead. Collectively, these studies suggest that arterial endothelial cells are specified and formed via repressing venous cell fate at the lateral plate mesoderm by Hh signaling during vasculogenesis.
Copyright 2010 Elsevier Inc. All rights reserved.
Figures
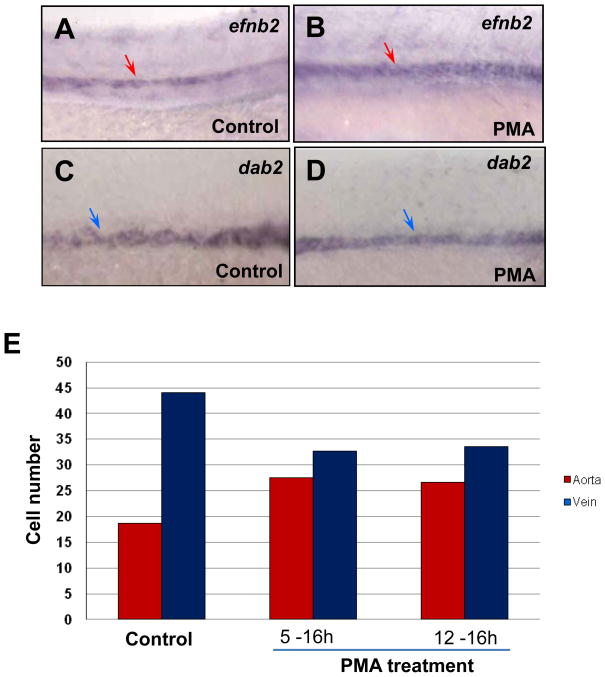
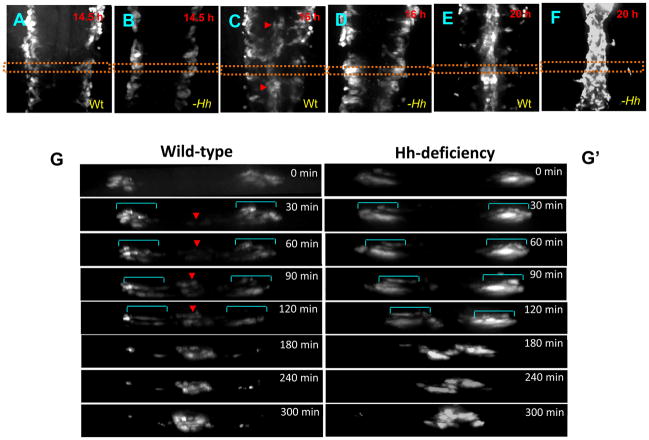
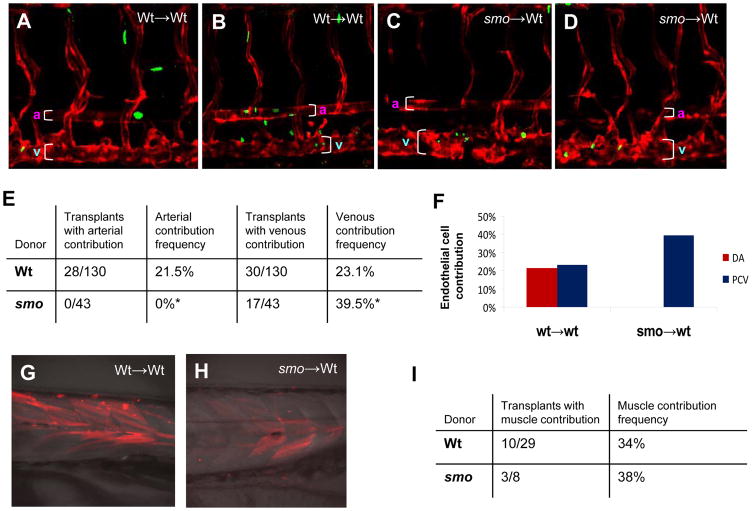
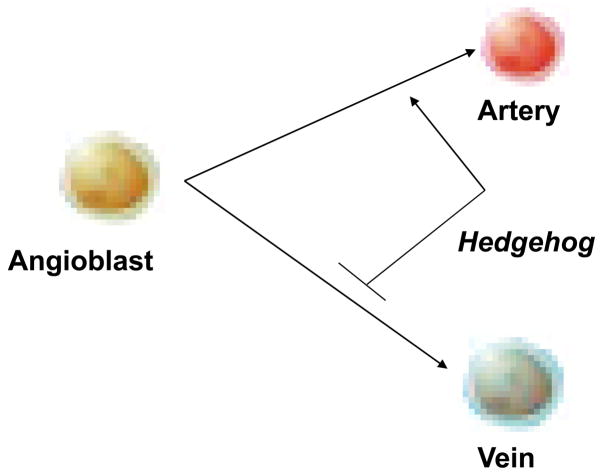
Similar articles
-
Hedgehog signaling via a calcitonin receptor-like receptor can induce arterial differentiation independently of VEGF signaling in zebrafish.Blood. 2012 Jul 12;120(2):477-88. doi: 10.1182/blood-2011-10-383729. Epub 2012 Jun 5. Blood. 2012. PMID: 22668851 Free PMC article.
-
Hedgehog signaling is required for differentiation of endocardial progenitors in zebrafish.Dev Biol. 2012 Jan 15;361(2):377-91. doi: 10.1016/j.ydbio.2011.11.004. Epub 2011 Nov 12. Dev Biol. 2012. PMID: 22119054
-
Arterial and venous progenitors of the major axial vessels originate at distinct locations.Dev Cell. 2013 Apr 29;25(2):196-206. doi: 10.1016/j.devcel.2013.03.017. Dev Cell. 2013. PMID: 23639444 Free PMC article.
-
Arterial-venous specification during development.Circ Res. 2009 Mar 13;104(5):576-88. doi: 10.1161/CIRCRESAHA.108.188805. Circ Res. 2009. PMID: 19286613 Review.
-
Specification of arterial, venous, and lymphatic endothelial cells during embryonic development.Histol Histopathol. 2010 May;25(5):637-46. doi: 10.14670/HH-25.637. Histol Histopathol. 2010. PMID: 20238301 Free PMC article. Review.
Cited by
-
Hedgehog signaling via a calcitonin receptor-like receptor can induce arterial differentiation independently of VEGF signaling in zebrafish.Blood. 2012 Jul 12;120(2):477-88. doi: 10.1182/blood-2011-10-383729. Epub 2012 Jun 5. Blood. 2012. PMID: 22668851 Free PMC article.
-
Nile Tilapia (Oreochromis niloticus) Patched1 Mutations Disrupt Cardiovascular Development and Vascular Integrity through Smoothened Signaling.Int J Mol Sci. 2024 Mar 15;25(6):3321. doi: 10.3390/ijms25063321. Int J Mol Sci. 2024. PMID: 38542295 Free PMC article.
-
Aplexone targets the HMG-CoA reductase pathway and differentially regulates arteriovenous angiogenesis.Development. 2011 Mar;138(6):1173-81. doi: 10.1242/dev.054049. Epub 2011 Feb 9. Development. 2011. PMID: 21307094 Free PMC article.
-
Todralazine protects zebrafish from lethal effects of ionizing radiation: role of hematopoietic cell expansion.Zebrafish. 2015 Feb;12(1):33-47. doi: 10.1089/zeb.2014.0992. Epub 2014 Dec 17. Zebrafish. 2015. PMID: 25517940 Free PMC article.
-
Arterial-venous network formation during brain vascularization involves hemodynamic regulation of chemokine signaling.Development. 2011 May;138(9):1717-26. doi: 10.1242/dev.059881. Epub 2011 Mar 23. Development. 2011. PMID: 21429983 Free PMC article.
References
-
- Appel B, Fritz A, Westerfield M, Grunwald DJ, Eisen JS, Riley BB. Delta-mediated specification of midline cell fates in zebrafish embryos. Curr Boil. 1999;9:247–56. - PubMed
-
- Barresi MJ, Stickney HL, Devoto SH. The zebrafish slow-muscle-omitted gene product is required for Hedgehog signal transduction and the development of slow muscle identity. Development. 2000;127:2189–99. - PubMed
-
- Bhat KM. The patched signaling pathway mediates repression of gooseberry allowing neuroblast specification by wingless during Drosophila neurogenesis. Development. 1996;122:2921–32. - PubMed
-
- Brown L, Rodaway A, Schilling T, Jowett T, Ingham P, Patient R, Sharrocks A. Insights into vasculogenesis revealed by expression of ETS-domain transcription factor Fli-1 in wild type and mutant zebrafish embryos. Mechanisms of development. 2000:237–252. - PubMed
-
- Byrd N, Beck S, Maye P, Narasimhaiah R, St-Jacques B, Zhang X, McMahon J, McMahon AP, Grabel L. Hedgehog is required fo murine yolk sac angiogenesis. Development. 2002;129:361–372. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
