

Dissecting electrostatic screening, specific ion binding, and ligand binding in an energetic model for glycine riboswitch folding
- PMID: 20194520
- PMCID: PMC2844619
- DOI: 10.1261/rna.1985110
Dissecting electrostatic screening, specific ion binding, and ligand binding in an energetic model for glycine riboswitch folding
Abstract
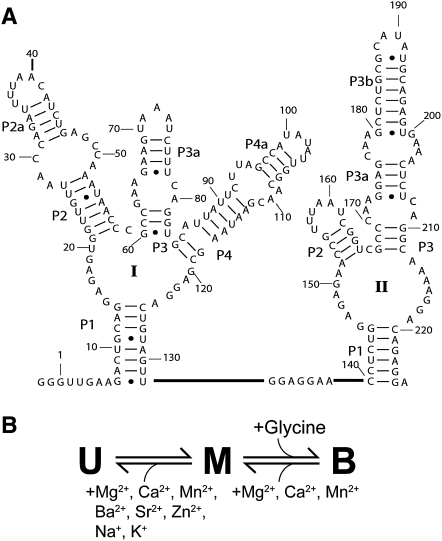
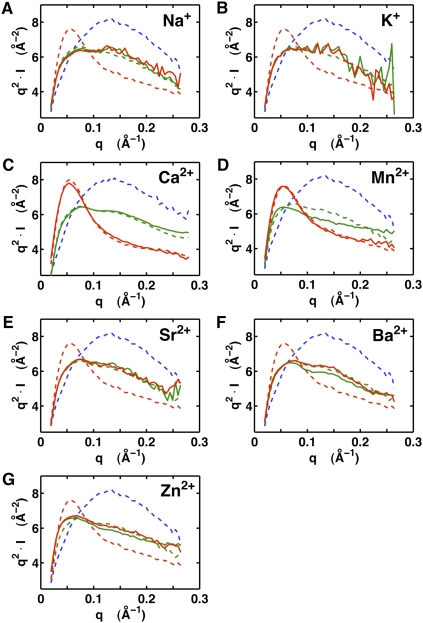
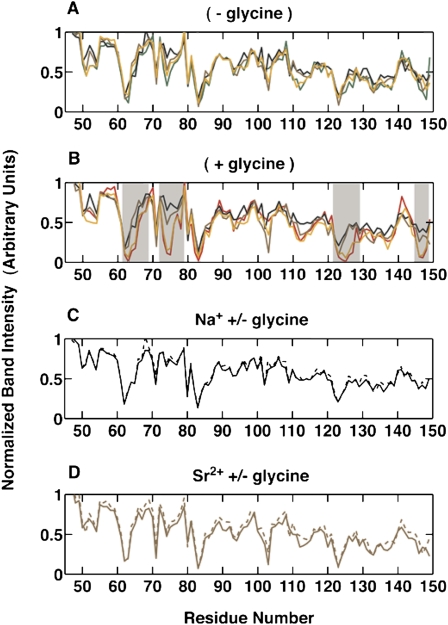
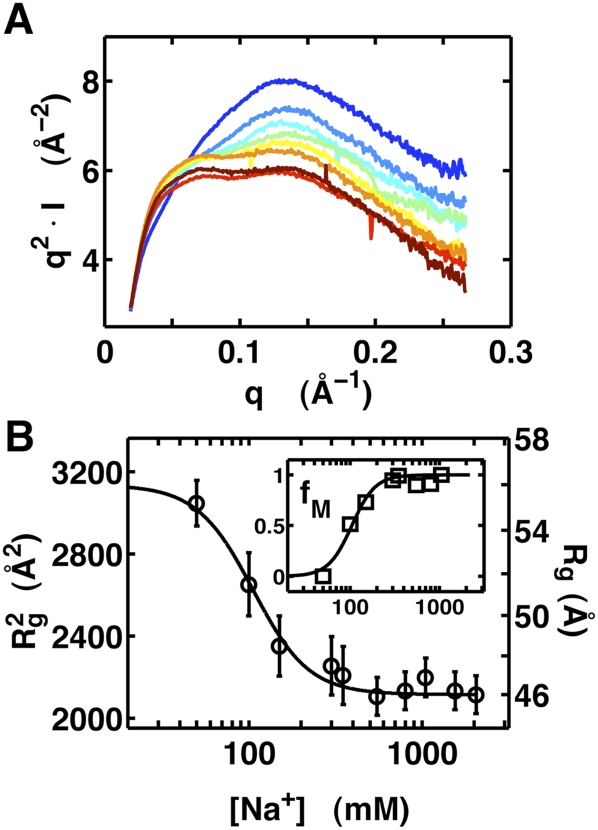
Riboswitches are gene-regulating RNAs that are usually found in the 5'-untranslated regions of messenger RNA. As the sugar-phosphate backbone of RNA is highly negatively charged, the folding and ligand-binding interactions of riboswitches are strongly dependent on the presence of cations. Using small angle X-ray scattering (SAXS) and hydroxyl radical footprinting, we examined the cation dependence of the different folding stages of the glycine-binding riboswitch from Vibrio cholerae. We found that the partial folding of the tandem aptamer of this riboswitch in the absence of glycine is supported by all tested mono- and divalent ions, suggesting that this transition is mediated by nonspecific electrostatic screening. Poisson-Boltzmann calculations using SAXS-derived low-resolution structural models allowed us to perform an energetic dissection of this process. The results showed that a model with a constant favorable contribution to folding that is opposed by an unfavorable electrostatic term that varies with ion concentration and valency provides a reasonable quantitative description of the observed folding behavior. Glycine binding, on the other hand, requires specific divalent ions binding based on the observation that Mg(2+), Ca(2+), and Mn(2+) facilitated glycine binding, whereas other divalent cations did not. The results provide a case study of how ion-dependent electrostatic relaxation, specific ion binding, and ligand binding can be coupled to shape the energetic landscape of a riboswitch and can begin to be quantitatively dissected.
Figures
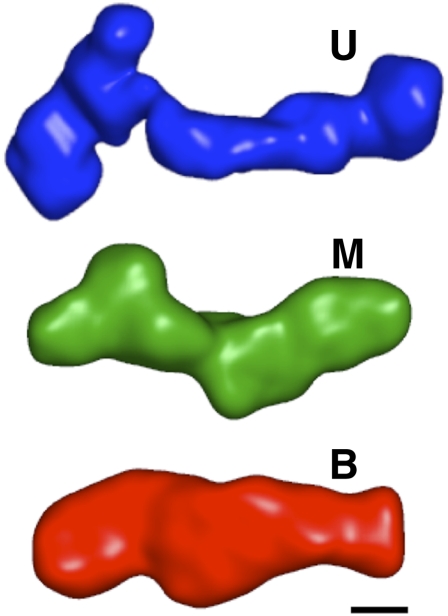
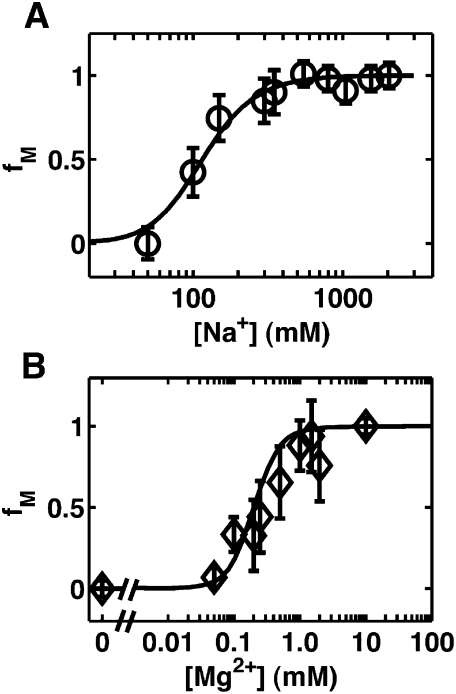
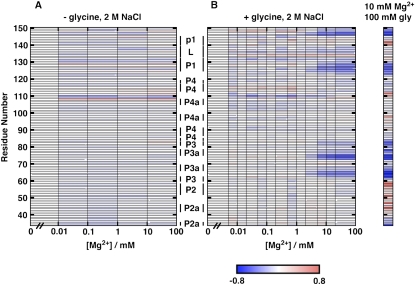
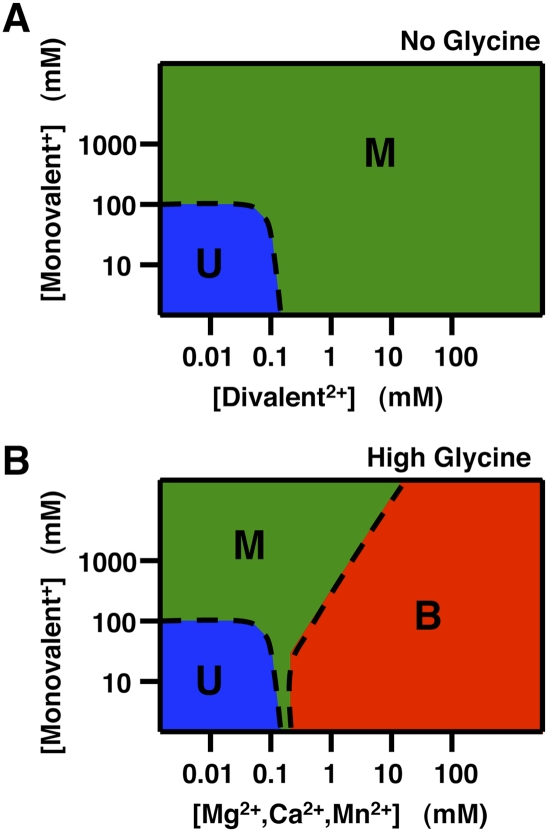
Similar articles
-
Riboswitch conformations revealed by small-angle X-ray scattering.Methods Mol Biol. 2009;540:141-59. doi: 10.1007/978-1-59745-558-9_11. Methods Mol Biol. 2009. PMID: 19381558 Free PMC article.
-
Structural transitions and thermodynamics of a glycine-dependent riboswitch from Vibrio cholerae.J Mol Biol. 2007 Feb 2;365(5):1393-406. doi: 10.1016/j.jmb.2006.10.022. Epub 2006 Oct 13. J Mol Biol. 2007. PMID: 17118400 Free PMC article.
-
Modulation of quaternary structure and enhancement of ligand binding by the K-turn of tandem glycine riboswitches.RNA. 2013 Feb;19(2):167-76. doi: 10.1261/rna.036269.112. Epub 2012 Dec 17. RNA. 2013. PMID: 23249744 Free PMC article.
-
Amino acid recognition and gene regulation by riboswitches.Biochim Biophys Acta. 2009 Sep-Oct;1789(9-10):592-611. doi: 10.1016/j.bbagrm.2009.07.002. Epub 2009 Jul 18. Biochim Biophys Acta. 2009. PMID: 19619684 Free PMC article. Review.
-
Purine sensing by riboswitches.Biol Cell. 2008 Jan;100(1):1-11. doi: 10.1042/BC20070088. Biol Cell. 2008. PMID: 18072940 Review.
Cited by
-
Predicting ion-nucleic acid interactions by energy landscape-guided sampling.J Chem Theory Comput. 2012 Jun 12;8(6):2095-2101. doi: 10.1021/ct300227a. Epub 2012 Apr 30. J Chem Theory Comput. 2012. PMID: 23002389 Free PMC article.
-
The expression platform and the aptamer: cooperativity between Mg2+ and ligand in the SAM-I riboswitch.Nucleic Acids Res. 2013 Feb 1;41(3):1922-35. doi: 10.1093/nar/gks978. Epub 2012 Dec 20. Nucleic Acids Res. 2013. PMID: 23258703 Free PMC article.
-
Increasing the length of poly-pyrimidine bulges broadens RNA conformational ensembles with minimal impact on stacking energetics.RNA. 2018 Oct;24(10):1363-1376. doi: 10.1261/rna.066258.118. Epub 2018 Jul 16. RNA. 2018. PMID: 30012568 Free PMC article.
-
Global analysis of riboswitches by small-angle X-ray scattering and calorimetry.Biochim Biophys Acta. 2014 Oct;1839(10):1020-1029. doi: 10.1016/j.bbagrm.2014.04.014. Epub 2014 Apr 24. Biochim Biophys Acta. 2014. PMID: 24769285 Free PMC article. Review.
-
Probing the kinetic and thermodynamic consequences of the tetraloop/tetraloop receptor monovalent ion-binding site in P4-P6 RNA by smFRET.Biochem Soc Trans. 2015 Apr;43(2):172-8. doi: 10.1042/BST20140268. Biochem Soc Trans. 2015. PMID: 25849913 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous