

The novel endosomal membrane protein Ema interacts with the class C Vps-HOPS complex to promote endosomal maturation
- PMID: 20194640
- PMCID: PMC2835942
- DOI: 10.1083/jcb.200911126
The novel endosomal membrane protein Ema interacts with the class C Vps-HOPS complex to promote endosomal maturation
Abstract
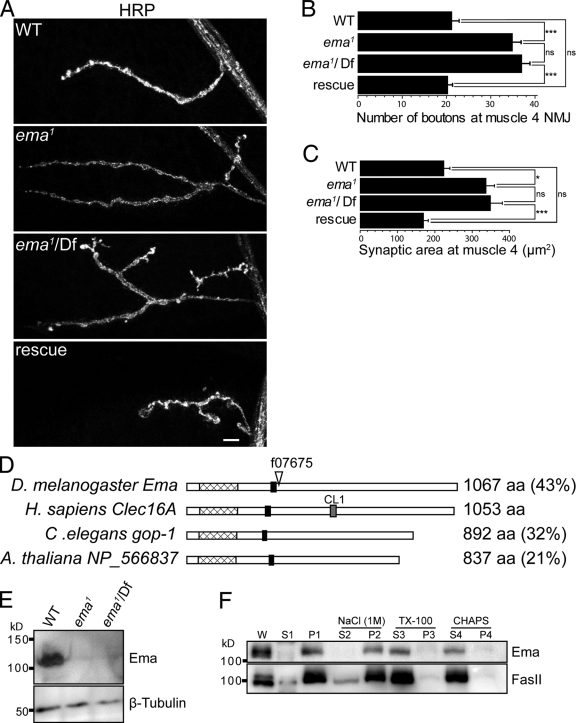
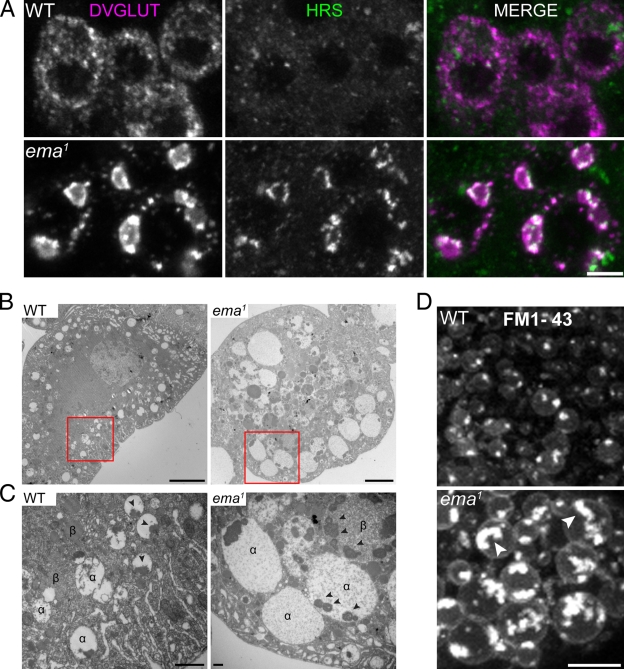
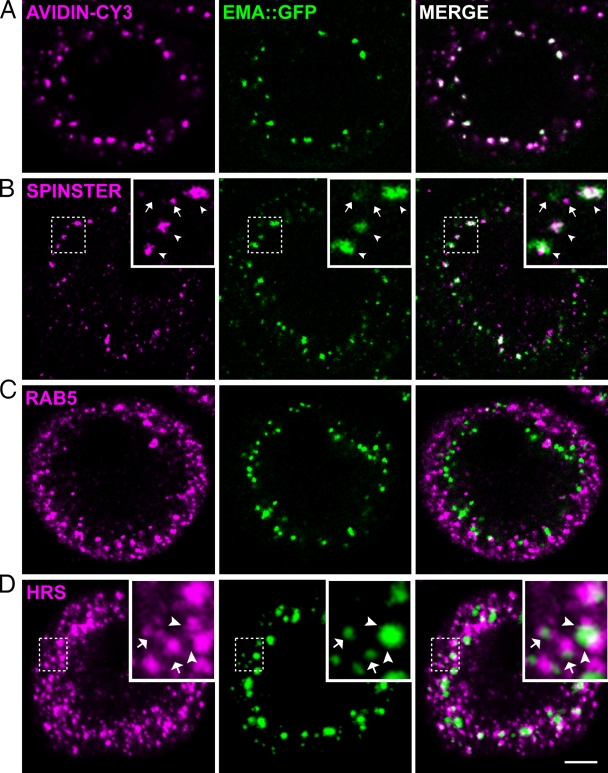
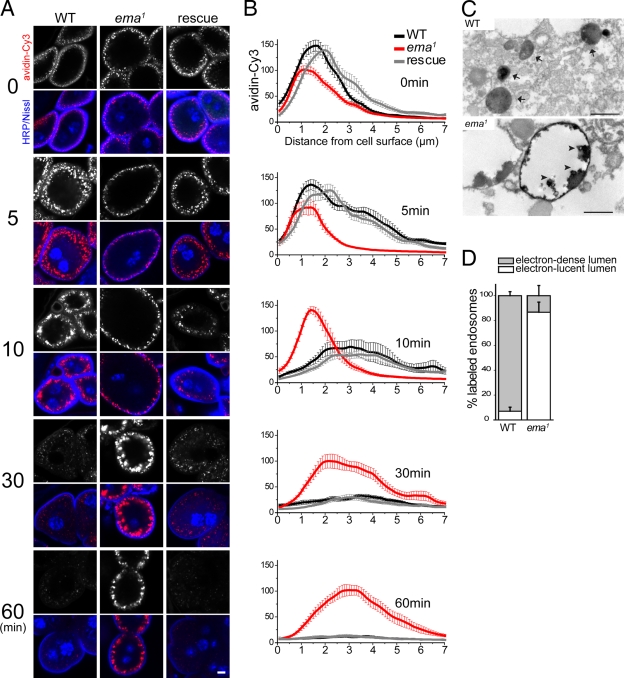
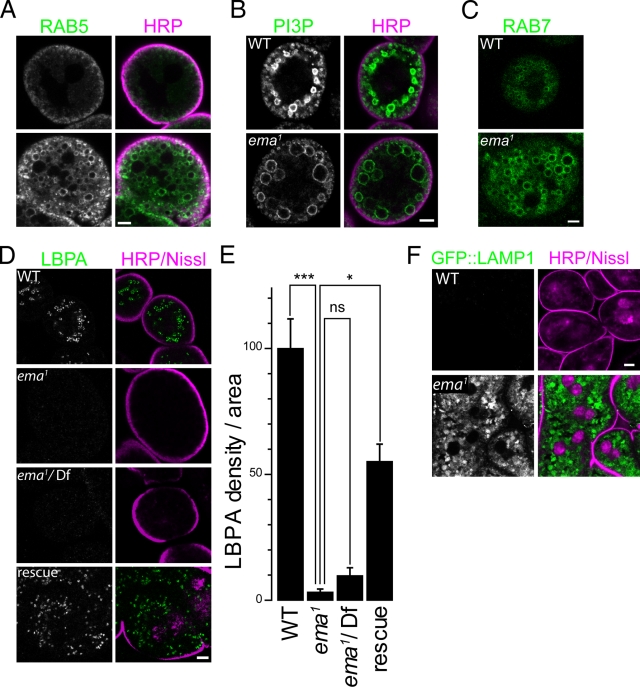
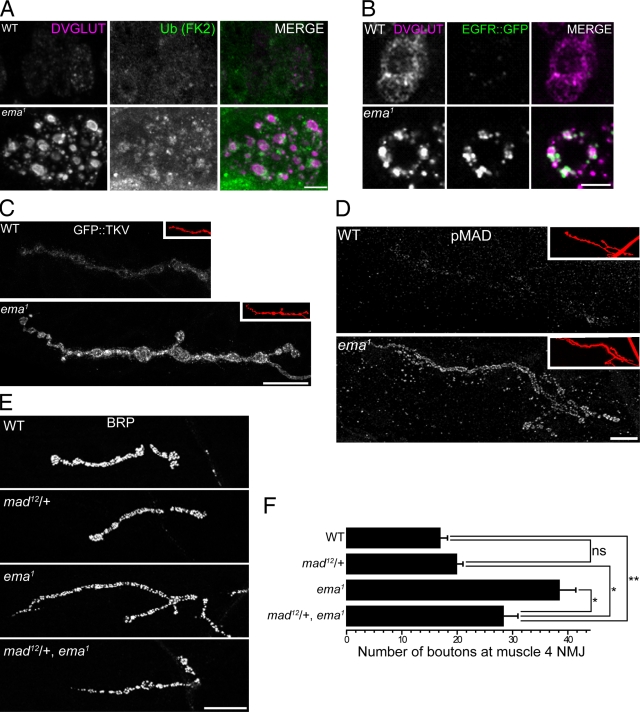
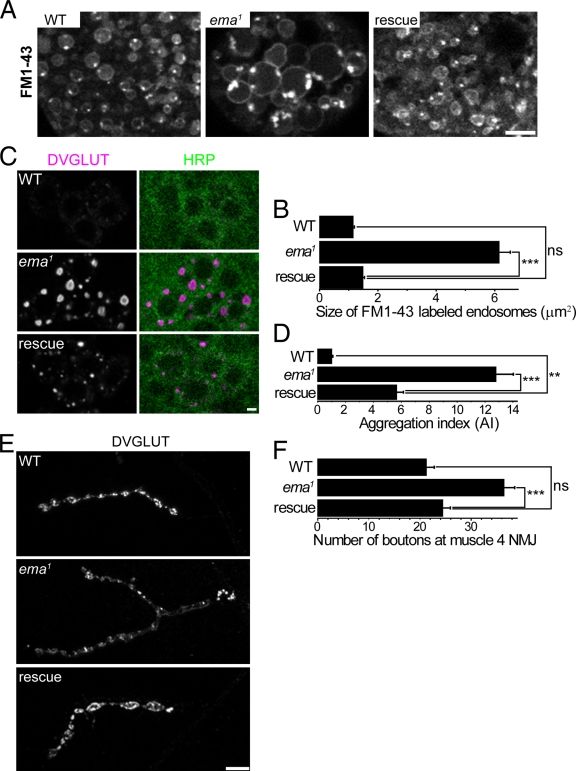
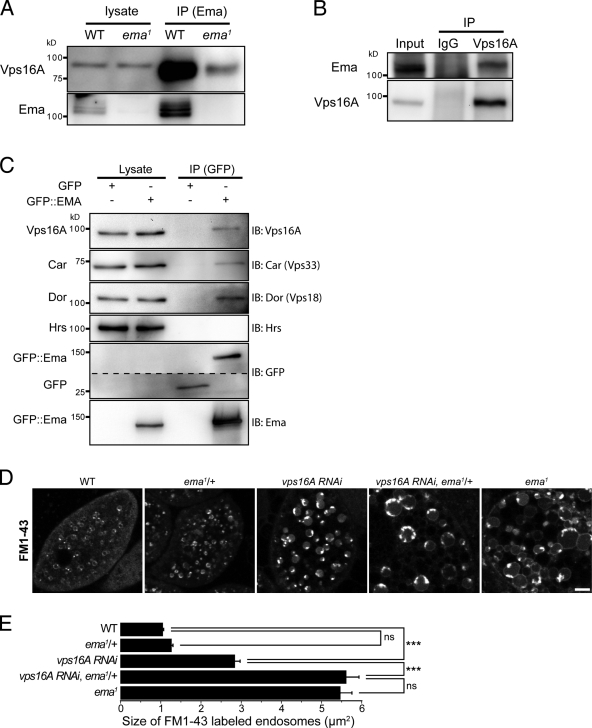
Endosomal maturation is critical for accurate and efficient cargo transport through endosomal compartments. Here we identify a mutation of the novel Drosophila gene, ema (endosomal maturation defective) in a screen for abnormal synaptic overgrowth and defective protein trafficking. Ema is an endosomal membrane protein required for trafficking of fluid-phase and receptor-mediated endocytic cargos. In the ema mutant, enlarged endosomal compartments accumulate as endosomal maturation fails, with early and late endosomes unable to progress into mature degradative late endosomes and lysosomes. Defective endosomal down-regulation of BMP signaling is responsible for the abnormal synaptic overgrowth. Ema binds to and genetically interacts with Vps16A, a component of the class C Vps-HOPS complex that promotes endosomal maturation. The human orthologue of ema, Clec16A, is a candidate susceptibility locus for autoimmune disorders, and its expression rescues the Drosophila mutant demonstrating conserved function. Characterizing this novel gene family identifies a new component of the endosomal pathway and provides insights into class C Vps-HOPS complex function.
Figures
References
-
- Awata T., Kawasaki E., Tanaka S., Ikegami H., Maruyama T., Shimada A., Nakanishi K., Kobayashi T., Iizuka H., Uga M., et al. ; Japanese Study Group on Type 1 Diabetes Genetics. 2009. Association of type 1 diabetes with two Loci on 12q13 and 16p13 and the influence coexisting thyroid autoimmunity in Japanese. J. Clin. Endocrinol. Metab. 94:231–235. 10.1210/jc.2008-0718 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
