

A ratchet mechanism for amplification in low-frequency mammalian hearing
- PMID: 20194771
- PMCID: PMC2841936
- DOI: 10.1073/pnas.0914345107
A ratchet mechanism for amplification in low-frequency mammalian hearing
Abstract
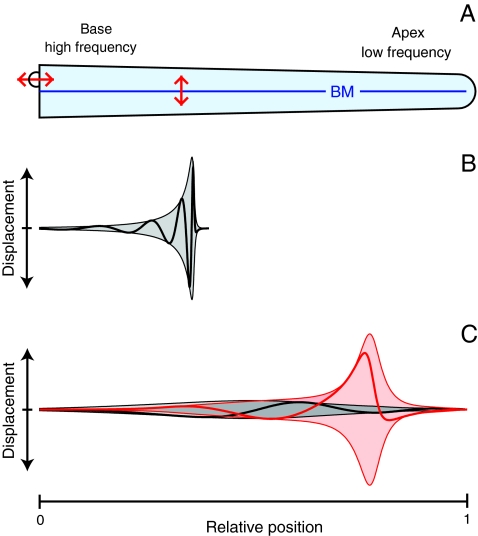
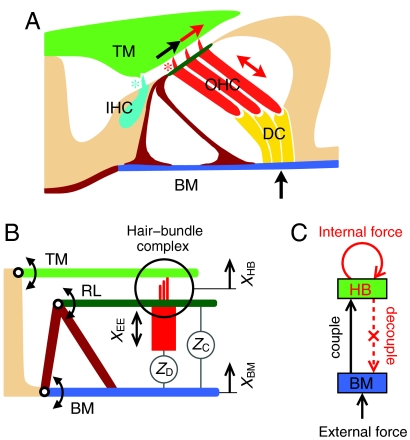
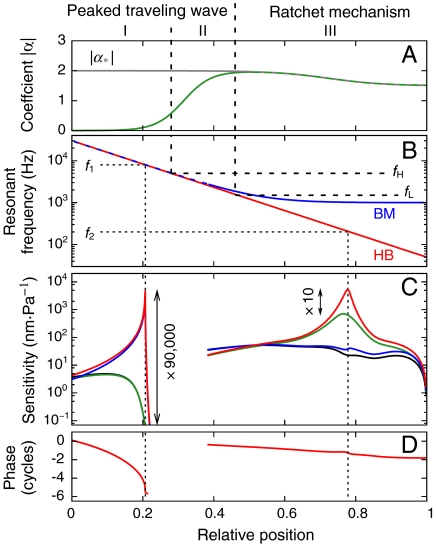
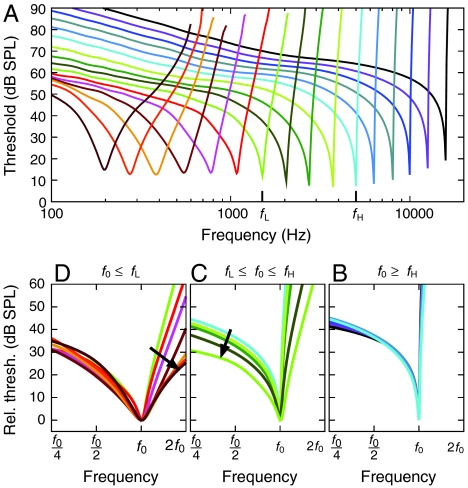
The sensitivity and frequency selectivity of hearing result from tuned amplification by an active process in the mechanoreceptive hair cells. In most vertebrates, the active process stems from the active motility of hair bundles. The mammalian cochlea exhibits an additional form of mechanical activity termed electromotility: its outer hair cells (OHCs) change length upon electrical stimulation. The relative contributions of these two mechanisms to the active process in the mammalian inner ear is the subject of intense current debate. Here, we show that active hair-bundle motility and electromotility can together implement an efficient mechanism for amplification that functions like a ratchet: Sound-evoked forces, acting on the basilar membrane, are transmitted to the hair bundles, whereas electromotility decouples active hair-bundle forces from the basilar membrane. This unidirectional coupling can extend the hearing range well below the resonant frequency of the basilar membrane. It thereby provides a concept for low-frequency hearing that accounts for a variety of unexplained experimental observations from the cochlear apex, including the shape and phase behavior of apical tuning curves, their lack of significant nonlinearities, and the shape changes of threshold tuning curves of auditory-nerve fibers along the cochlea. The ratchet mechanism constitutes a general design principle for implementing mechanical amplification in engineering applications.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Amplification and Suppression of Traveling Waves along the Mouse Organ of Corti: Evidence for Spatial Variation in the Longitudinal Coupling of Outer Hair Cell-Generated Forces.J Neurosci. 2019 Mar 6;39(10):1805-1816. doi: 10.1523/JNEUROSCI.2608-18.2019. Epub 2019 Jan 16. J Neurosci. 2019. PMID: 30651330 Free PMC article.
-
Vibration of the organ of Corti within the cochlear apex in mice.J Neurophysiol. 2014 Sep 1;112(5):1192-204. doi: 10.1152/jn.00306.2014. Epub 2014 Jun 11. J Neurophysiol. 2014. PMID: 24920025 Free PMC article.
-
Static length changes of cochlear outer hair cells can tune low-frequency hearing.PLoS Comput Biol. 2018 Jan 19;14(1):e1005936. doi: 10.1371/journal.pcbi.1005936. eCollection 2018 Jan. PLoS Comput Biol. 2018. PMID: 29351276 Free PMC article.
-
The cortilymph wave: Its relation to the traveling wave, auditory-nerve responses, and low-frequency downward glides.Hear Res. 2025 Jun;462:109279. doi: 10.1016/j.heares.2025.109279. Epub 2025 Apr 16. Hear Res. 2025. PMID: 40253777 Review.
-
Tuned hair cells for hearing, but tuned basilar membrane for overload protection: evidence from dolphins, bats, and desert rodents.Hear Res. 1994 Jul;78(1):98-114. doi: 10.1016/0378-5955(94)90048-5. Hear Res. 1994. PMID: 7961182 Review.
Cited by
-
Contribution of active hair-bundle motility to nonlinear amplification in the mammalian cochlea.Proc Natl Acad Sci U S A. 2012 Dec 18;109(51):21076-80. doi: 10.1073/pnas.1219379110. Epub 2012 Dec 3. Proc Natl Acad Sci U S A. 2012. PMID: 23213236 Free PMC article.
-
Effect of Contralateral Medial Olivocochlear Feedback on Perceptual Estimates of Cochlear Gain and Compression.J Assoc Res Otolaryngol. 2016 Dec;17(6):559-575. doi: 10.1007/s10162-016-0574-8. Epub 2016 Aug 22. J Assoc Res Otolaryngol. 2016. PMID: 27550069 Free PMC article. Clinical Trial.
-
Coupling active hair bundle mechanics, fast adaptation, and somatic motility in a cochlear model.Biophys J. 2011 Jun 8;100(11):2576-85. doi: 10.1016/j.bpj.2011.04.049. Biophys J. 2011. PMID: 21641302 Free PMC article.
-
A critique of the critical cochlea: Hopf--a bifurcation--is better than none.J Neurophysiol. 2010 Sep;104(3):1219-29. doi: 10.1152/jn.00437.2010. Epub 2010 Jun 10. J Neurophysiol. 2010. PMID: 20538769 Free PMC article. Review.
-
Reticular lamina and basilar membrane vibrations in living mouse cochleae.Proc Natl Acad Sci U S A. 2016 Aug 30;113(35):9910-5. doi: 10.1073/pnas.1607428113. Epub 2016 Aug 11. Proc Natl Acad Sci U S A. 2016. PMID: 27516544 Free PMC article.
References
-
- Lighthill J. Energy flow in the cochlea. J Fluid Mech. 1981;106:149–213.
-
- Mammano F, Nobili R. Biophysics of the cochlea: Linear approximation. J Acoust Soc Am. 1993;93:3320–3332. - PubMed
-
- Siebert WM. Stimulus transformations in the peripheral auditory system. In: Kolers PA, Eden M, editors. Recognizing Patterns. Cambridge: MIT; 1968. pp. 104–133.
-
- Zweig G. In Cold Spring Harbor Symp Quan Biol. Vol. 40. Cold Spring Harbor Laboratory; 1976. Basilar membrane motion; pp. 619–633. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
