

A tomato strigolactone-impaired mutant displays aberrant shoot morphology and plant interactions
- PMID: 20194924
- PMCID: PMC2852664
- DOI: 10.1093/jxb/erq041
A tomato strigolactone-impaired mutant displays aberrant shoot morphology and plant interactions
Abstract
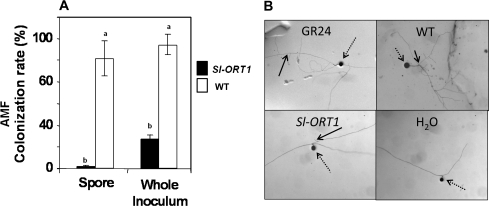
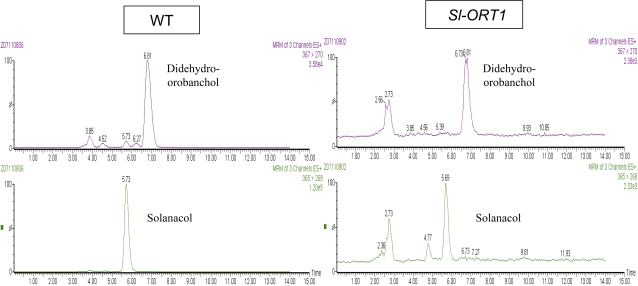
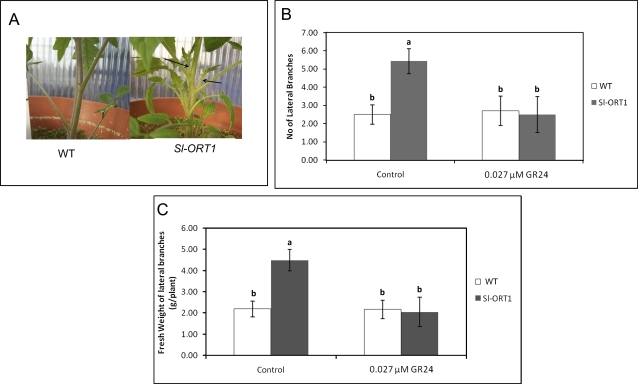
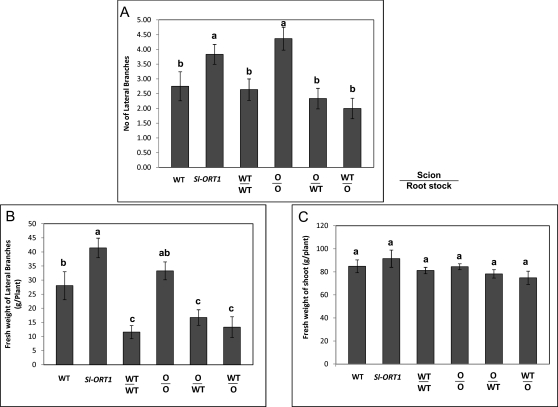
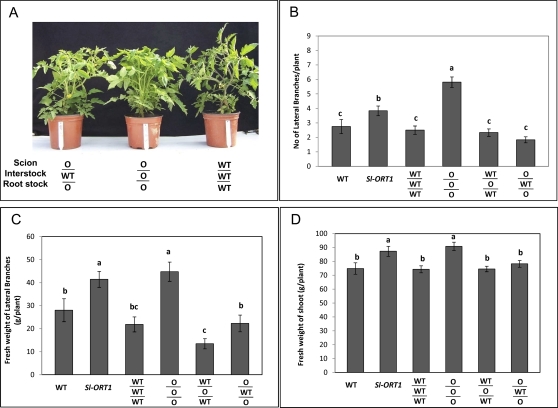
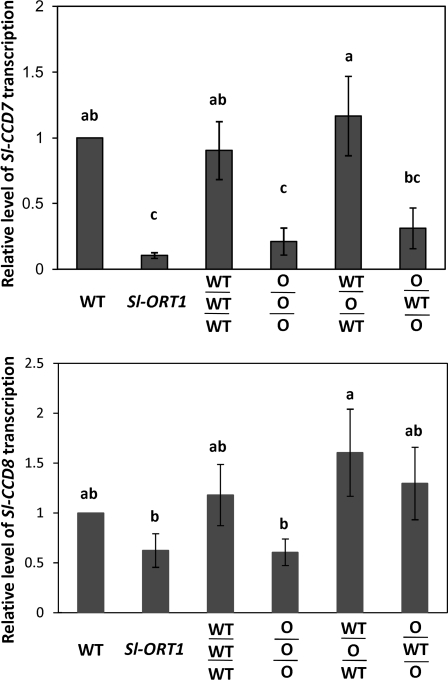
Strigolactones are considered a new group of plant hormones. Their role as modulators of plant growth and signalling molecules for plant interactions first became evident in Arabidopsis, pea, and rice mutants that were flawed in strigolactone production, release, or perception. The first evidence in tomato (Solanum lycopersicon) of strigolactone deficiency is presented here. Sl-ORT1, previously identified as resistant to the parasitic plant Orobanche, had lower levels of arbuscular mycorrhizal fungus (Glomus intraradices) colonization, possibly as a result of its reduced ability to induce mycorrhizal hyphal branching. Biochemical analysis of mutant root extracts suggested that it produces only minute amounts of two of the tomato strigolactones: solanacol and didehydro-orobanchol. Accordingly, the transcription level of a key enzyme (CCD7) putatively involved in strigolactone synthesis in tomato was reduced in Sl-ORT1 compared with the wild type (WT). Sl-ORT1 shoots exhibited increased lateral shoot branching, whereas exogenous application of the synthetic strigolactone GR24 to the mutant restored the WT phenotype by reducing the number of lateral branches. Reduced lateral shoot branching was also evident in grafted plants which included a WT interstock, which was grafted between the mutant rootstock and the scion. In roots of these grafted plants, the CCD7 transcription level was not significantly induced, nor was mycorrhizal sensitivity restored. Hence, WT-interstock grafting, which restores mutant shoot morphology to WT, does not restore mutant root properties to WT. Characterization of the first tomato strigolactone-deficient mutant supports the putative general role of strigolactones as messengers of suppression of lateral shoot branching in a diversity of plant species.
Figures
Similar articles
-
SlCCD7 controls strigolactone biosynthesis, shoot branching and mycorrhiza-induced apocarotenoid formation in tomato.Plant J. 2010 Jan;61(2):300-11. doi: 10.1111/j.1365-313X.2009.04056.x. Epub 2009 Oct 20. Plant J. 2010. PMID: 19845881
-
The tomato MAX1 homolog, SlMAX1, is involved in the biosynthesis of tomato strigolactones from carlactone.New Phytol. 2018 Jul;219(1):297-309. doi: 10.1111/nph.15131. Epub 2018 Apr 14. New Phytol. 2018. PMID: 29655242
-
Strigolactone deficiency confers resistance in tomato line SL-ORT1 to the parasitic weeds Phelipanche and Orobanche spp.Phytopathology. 2011 Feb;101(2):213-22. doi: 10.1094/PHYTO-07-10-0184. Phytopathology. 2011. PMID: 20942651
-
Strigolactones: structures and biological activities.Pest Manag Sci. 2009 May;65(5):467-70. doi: 10.1002/ps.1726. Pest Manag Sci. 2009. PMID: 19222028 Review.
-
New genes in the strigolactone-related shoot branching pathway.Curr Opin Plant Biol. 2010 Feb;13(1):34-9. doi: 10.1016/j.pbi.2009.10.003. Epub 2009 Nov 11. Curr Opin Plant Biol. 2010. PMID: 19913454 Review.
Cited by
-
Early transcriptional defense responses in Arabidopsis cell suspension culture under high-light conditions.Plant Physiol. 2011 Jul;156(3):1439-56. doi: 10.1104/pp.111.177766. Epub 2011 Apr 29. Plant Physiol. 2011. PMID: 21531897 Free PMC article.
-
Strigolactone involvement in root development, response to abiotic stress, and interactions with the biotic soil environment.Plant Physiol. 2014 Oct;166(2):560-9. doi: 10.1104/pp.114.244939. Epub 2014 Jul 18. Plant Physiol. 2014. PMID: 25037210 Free PMC article.
-
ZmCCD7/ZpCCD7 encodes a carotenoid cleavage dioxygenase mediating shoot branching.Planta. 2016 Jun;243(6):1407-18. doi: 10.1007/s00425-016-2479-5. Epub 2016 Feb 19. Planta. 2016. PMID: 26895334
-
The role of plant hormones during grafting.J Plant Res. 2018 Jan;131(1):49-58. doi: 10.1007/s10265-017-0994-5. Epub 2017 Nov 27. J Plant Res. 2018. PMID: 29181647 Free PMC article. Review.
-
Strigolactones and Cytokinin Interaction in Buds in the Control of Rice Tillering.Front Plant Sci. 2022 Jul 1;13:837136. doi: 10.3389/fpls.2022.837136. eCollection 2022. Front Plant Sci. 2022. PMID: 35845690 Free PMC article.
References
-
- Akiyama K, Matsuzaki K, Hayashi H. Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature. 2005;435:824–827. - PubMed
-
- Bainbridge K, Sorefan K, Ward S, Leyser O. Hormonally controlled expression of the Arabidopsis MAX4 shoot branching regulatory gene. The Plant Journal. 2005;44:569–580. - PubMed
-
- Bennett T, Sieberer T, Willett B, Booker J, Luschnig C, Leyser O. The Arabidopsis MAX pathway controls shoot branching by regulating auxin transport. Current Biology. 2006;16:553–563. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
