

Decoupling Environment-Dependent and Independent Genetic Robustness across Bacterial Species
- PMID: 20195496
- PMCID: PMC2829043
- DOI: 10.1371/journal.pcbi.1000690
Decoupling Environment-Dependent and Independent Genetic Robustness across Bacterial Species
Abstract
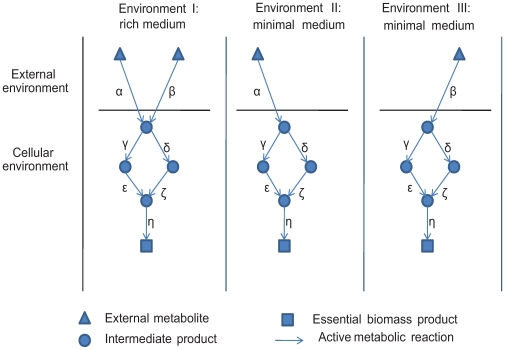
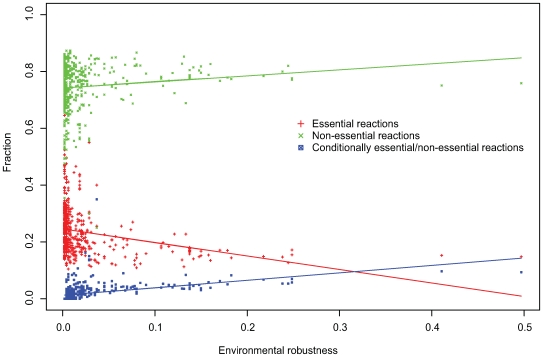
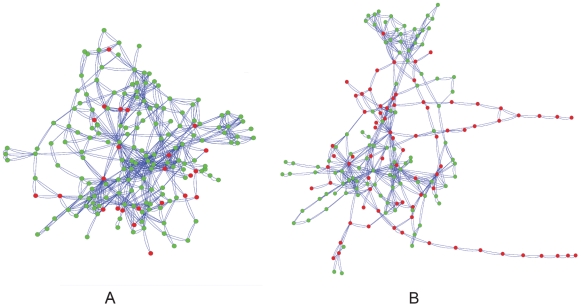
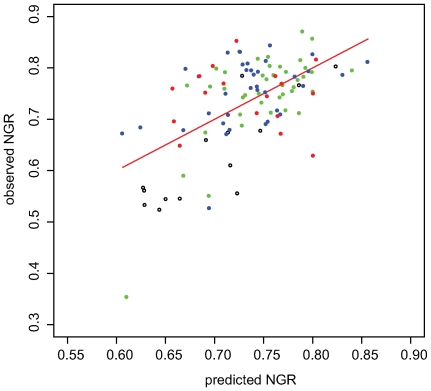
The evolutionary origins of genetic robustness are still under debate: it may arise as a consequence of requirements imposed by varying environmental conditions, due to intrinsic factors such as metabolic requirements, or directly due to an adaptive selection in favor of genes that allow a species to endure genetic perturbations. Stratifying the individual effects of each origin requires one to study the pertaining evolutionary forces across many species under diverse conditions. Here we conduct the first large-scale computational study charting the level of robustness of metabolic networks of hundreds of bacterial species across many simulated growth environments. We provide evidence that variations among species in their level of robustness reflect ecological adaptations. We decouple metabolic robustness into two components and quantify the extents of each: the first, environmental-dependent, is responsible for at least 20% of the non-essential reactions and its extent is associated with the species' lifestyle (specialized/generalist); the second, environmental-independent, is associated (correlation = approximately 0.6) with the intrinsic metabolic capacities of a species-higher robustness is observed in fast growers or in organisms with an extensive production of secondary metabolites. Finally, we identify reactions that are uniquely susceptible to perturbations in human pathogens, potentially serving as novel drug-targets.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- de Visser JA, Hermisson J, Wagner GP, Ancel Meyers L, Bagheri-Chaichian H, et al. Perspective: Evolution and detection of genetic robustness. Evolution. 2003;57:1959–1972. - PubMed
-
- Wagner A. Distributed robustness versus redundancy as causes of mutational robustness. Bioessays. 2005;27:176–188. - PubMed
-
- Kitano H. Biological robustness. Nat Rev Genet. 2004;5:826–837. - PubMed
-
- Pal C, Papp B, Lercher MJ. Adaptive evolution of bacterial metabolic networks by horizontal gene transfer. Nat Genet. 2005;37:1372–1375. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
