

Mycobacterium tuberculosis peptides presented by HLA-E molecules are targets for human CD8 T-cells with cytotoxic as well as regulatory activity
- PMID: 20195504
- PMCID: PMC2829052
- DOI: 10.1371/journal.ppat.1000782
Mycobacterium tuberculosis peptides presented by HLA-E molecules are targets for human CD8 T-cells with cytotoxic as well as regulatory activity
Abstract
Tuberculosis (TB) is an escalating global health problem and improved vaccines against TB are urgently needed. HLA-E restricted responses may be of interest for vaccine development since HLA-E displays very limited polymorphism (only 2 coding variants exist), and is not down-regulated by HIV-infection. The peptides from Mycobacterium tuberculosis (Mtb) potentially presented by HLA-E molecules, however, are unknown. Here we describe human T-cell responses to Mtb-derived peptides containing predicted HLA-E binding motifs and binding-affinity for HLA-E. We observed CD8(+) T-cell proliferation to the majority of the 69 peptides tested in Mtb responsive adults as well as in BCG-vaccinated infants. CD8(+) T-cells were cytotoxic against target-cells transfected with HLA-E only in the presence of specific peptide. These T cells were also able to lyse M. bovis BCG infected, but not control monocytes, suggesting recognition of antigens during mycobacterial infection. In addition, peptide induced CD8(+) T-cells also displayed regulatory activity, since they inhibited T-cell proliferation. This regulatory activity was cell contact-dependent, and at least partly dependent on membrane-bound TGF-beta. Our results significantly increase our understanding of the human immune response to Mtb by identification of CD8(+) T-cell responses to novel HLA-E binding peptides of Mtb, which have cytotoxic as well as immunoregulatory activity.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
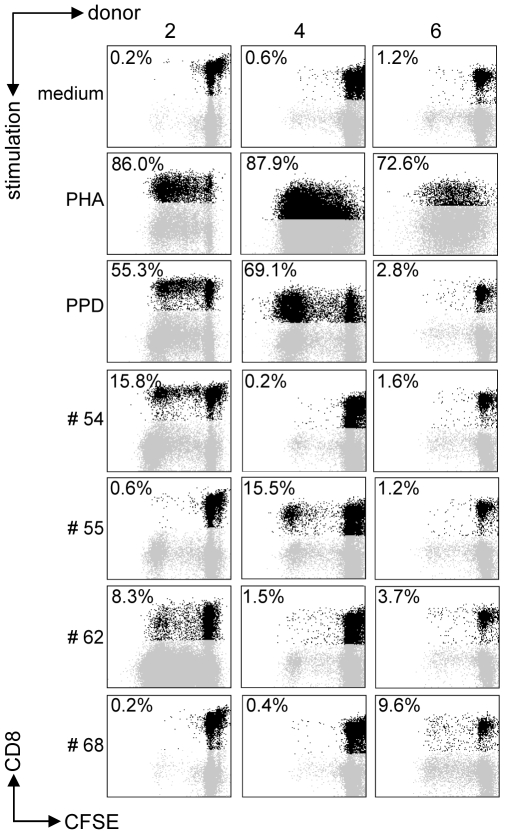
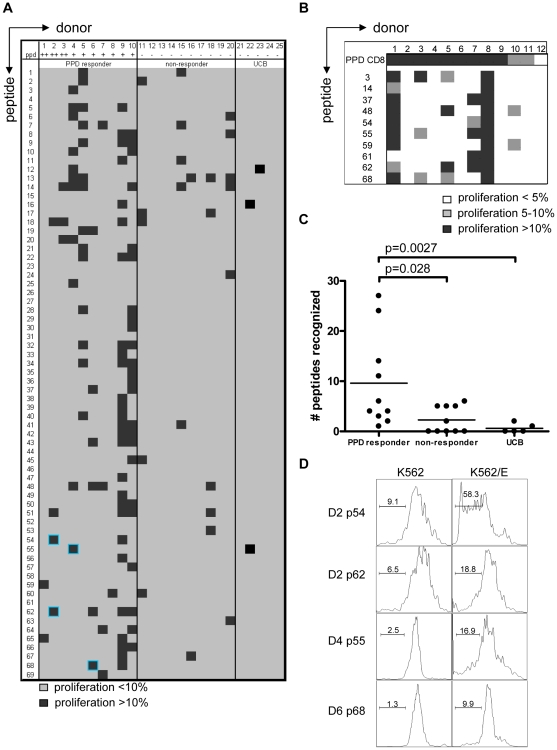
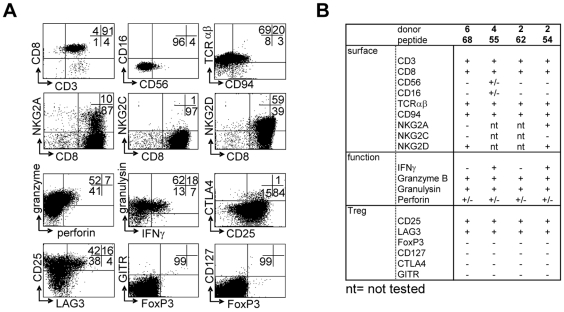
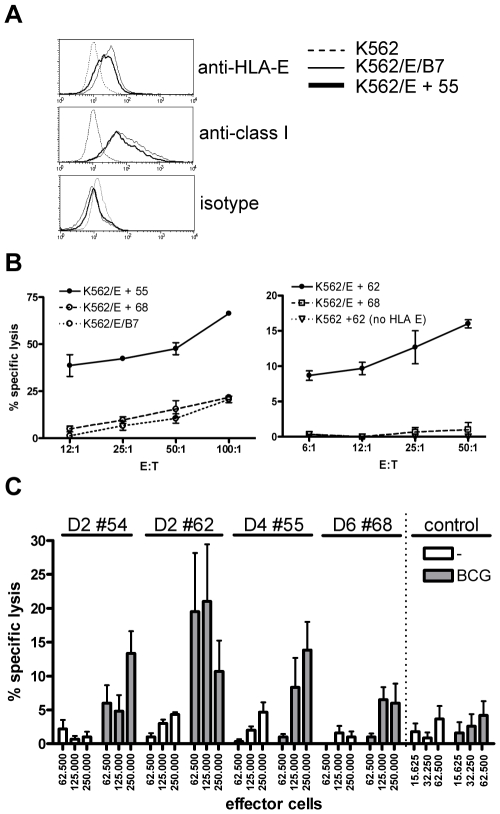
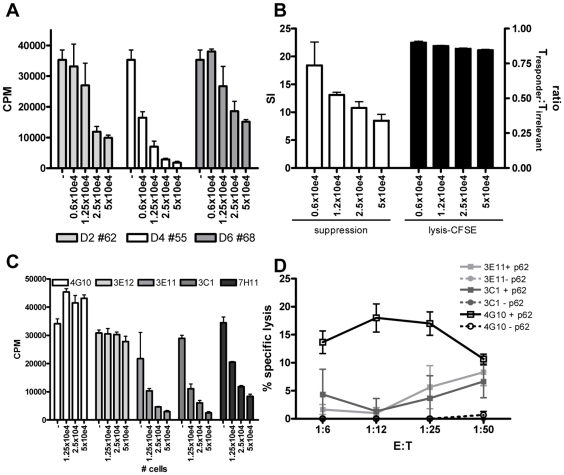
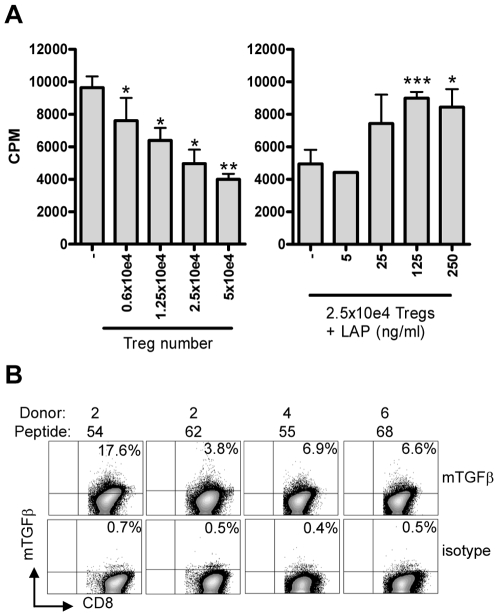
References
-
- World Health Organization. Global Tuberculosis Control. 2004. WHO report, WHO, Geneva.
-
- Fine PE, Rodrigues LC. Modern vaccines. Mycobacterial diseases. Lancet. 1990;335:1016–1020. - PubMed
-
- Ottenhoff TH. Overcoming the global crisis: “yes, we can”, but also for TB …? Eur J Immunol. 2009;39:2014–2020. - PubMed
-
- Kaufmann SH. Envisioning future strategies for vaccination against tuberculosis. Nat Rev Immunol. 2006;6:699–704. - PubMed
-
- Kaufmann SH. How can immunology contribute to the control of tuberculosis? Nat Rev Immunol. 2001;1:20–30. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
