

miR-24 regulates apoptosis by targeting the open reading frame (ORF) region of FAF1 in cancer cells
- PMID: 20195546
- PMCID: PMC2828487
- DOI: 10.1371/journal.pone.0009429
miR-24 regulates apoptosis by targeting the open reading frame (ORF) region of FAF1 in cancer cells
Abstract
Background: microRNAs (miRNAs) are small noncoding RNAs that regulate cognate mRNAs at the post-transcriptional stage. Several studies have shown that miRNAs modulate gene expression in mammalian cells by base pairing to complementary sites in the 3'-untranslated region (3'-UTR) of the target mRNAs.
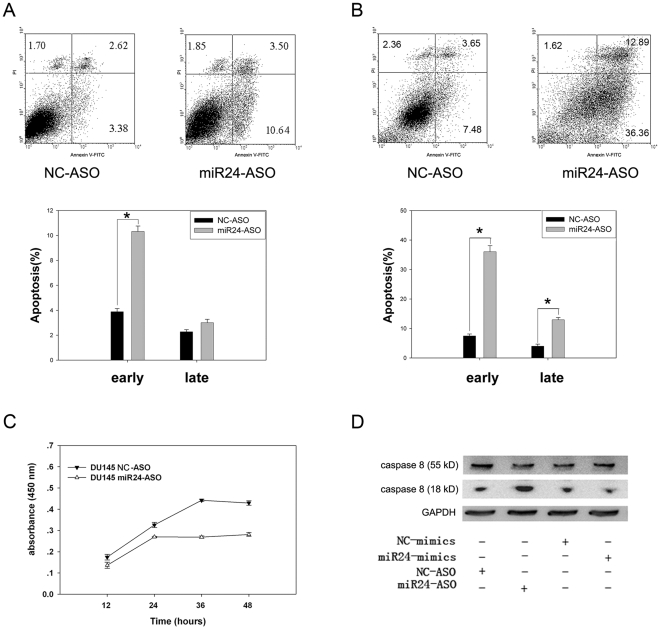
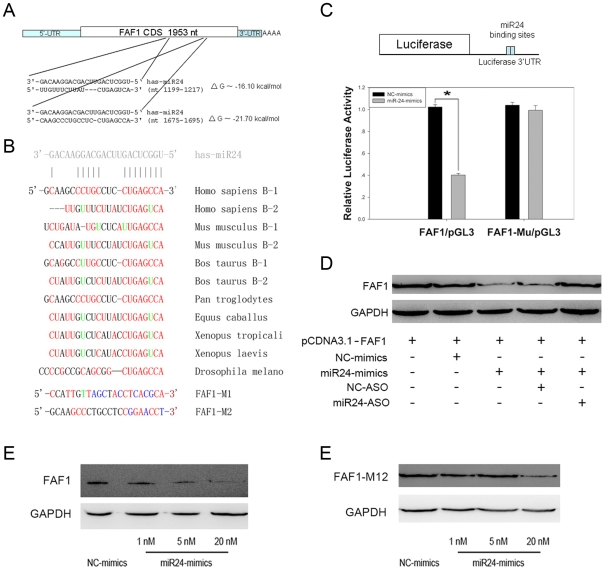
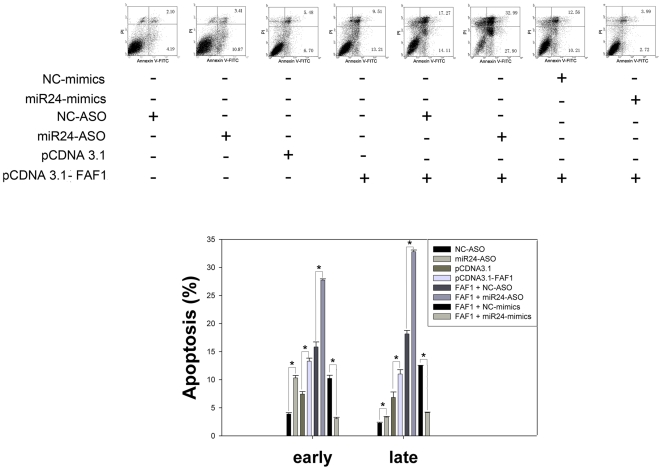
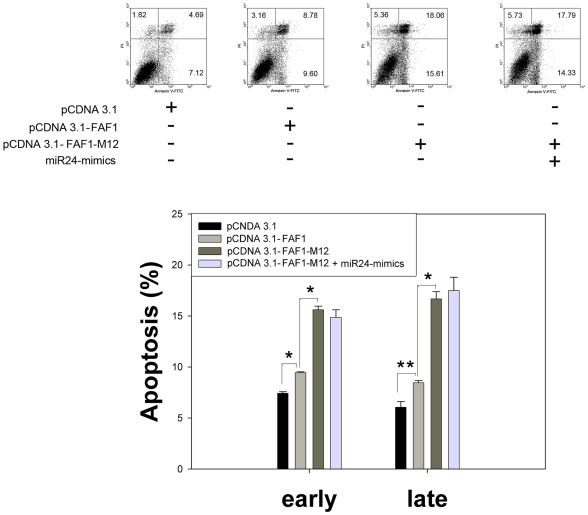
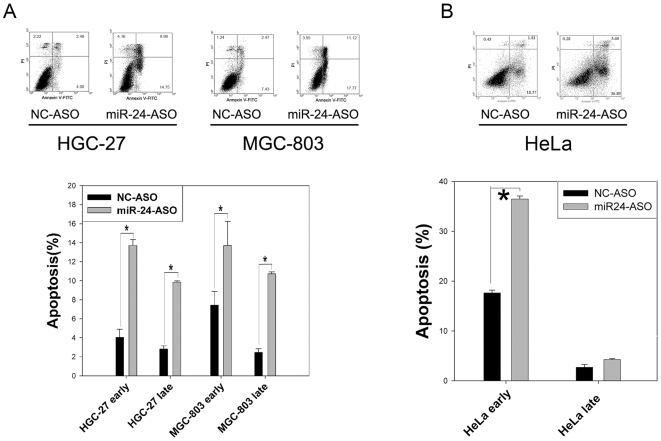
Methodology/principal findings: In the present study, miR-24 was found to target fas associated factor 1(FAF1) by binding to its amino acid coding sequence (CDS) region, thereby regulating apoptosis in DU-145 cells. This result supports an augmented model whereby animal miRNAs can exercise their effects through binding to the CDS region of the target mRNA. Transfection of miR-24 antisense oligonucleotide (miR-24-ASO) also induced apoptosis in HGC-27, MGC-803 and HeLa cells.
Conclusions/significance: We found that miR-24 regulates apoptosis by targeting FAF1 in cancer cells. These findings suggest that miR-24 could be an effective drug target for treatment of hormone-insensitive prostate cancer or other types of cancers. Future work may further develop miR-24 for therapeutic applications in cancer biology.
Conflict of interest statement
Figures
References
-
- Axtell MJ. Evolution of microRNAs and their targets: are all microRNAs biologically relevant? Biochim Biophys Acta. 2008;1779:725–734. - PubMed
-
- Yekta S, Shih IH, Bartel DP. MicroRNA-directed cleavage of HOXB8 mRNA. Science. 2004;304:594–596. - PubMed
-
- Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116:281–297. - PubMed
-
- Alvarez-Garcia IM, EA. MicroRNA functions in animal development and human disease. Development. 2005;132:10. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
