

Domain architecture evolution of pattern-recognition receptors
- PMID: 20195594
- PMCID: PMC2858798
- DOI: 10.1007/s00251-010-0428-1
Domain architecture evolution of pattern-recognition receptors
Abstract
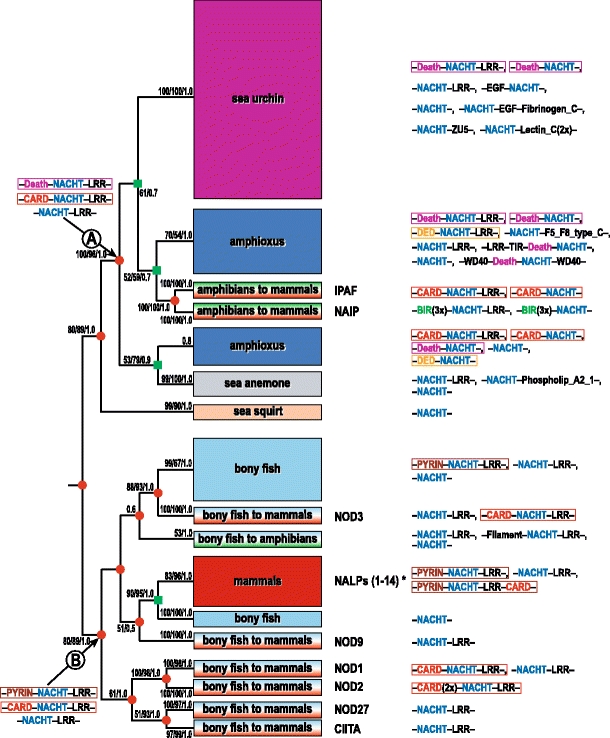
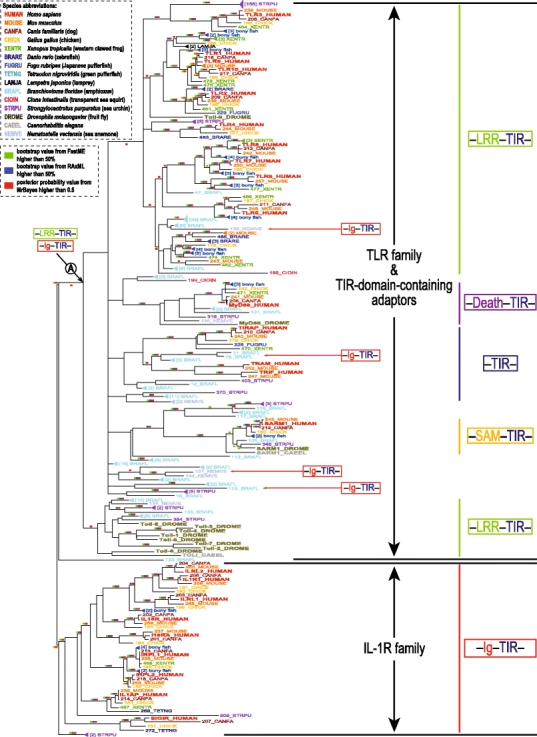
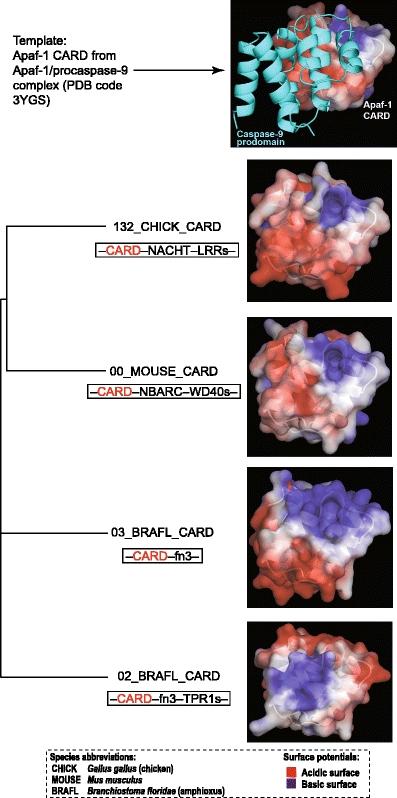
In animals, the innate immune system is the first line of defense against invading microorganisms, and the pattern-recognition receptors (PRRs) are the key components of this system, detecting microbial invasion and initiating innate immune defenses. Two families of PRRs, the intracellular NOD-like receptors (NLRs) and the transmembrane Toll-like receptors (TLRs), are of particular interest because of their roles in a number of diseases. Understanding the evolutionary history of these families and their pattern of evolutionary changes may lead to new insights into the functioning of this critical system. We found that the evolution of both NLR and TLR families included massive species-specific expansions and domain shuffling in various lineages, which resulted in the same domain architectures evolving independently within different lineages in a process that fits the definition of parallel evolution. This observation illustrates both the dynamics of the innate immune system and the effects of "combinatorially constrained" evolution, where existence of the limited numbers of functionally relevant domains constrains the choices of domain architectures for new members in the family, resulting in the emergence of independently evolved proteins with identical domain architectures, often mistaken for orthologs.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
