

Review
doi: 10.1007/s00018-009-0192-2.
Progress in phage display: evolution of the technique and its application
Affiliations
- PMID: 20196239
- PMCID: PMC11115567
- DOI: 10.1007/s00018-009-0192-2
Item in Clipboard
Review
Progress in phage display: evolution of the technique and its application
Cell Mol Life Sci.
2010 Mar.
Abstract
Phage display, the presentation of (poly)peptides as fusions to capsid proteins on the surface of bacterial viruses, celebrates its 25th birthday in 2010. The technique, coupled with in vitro selection, enables rapid identification and optimization of proteins based on their structural or functional properties. In the last two decades, it has advanced tremendously and has become widely accepted by the scientific community. This by no means exhaustive review aims to inform the reader of the key modifications in phage display. Novel display formats, innovative library designs and screening strategies are discussed. I will also briefly review some recent uses of the technology to illustrate its incredible versatility.
Figures
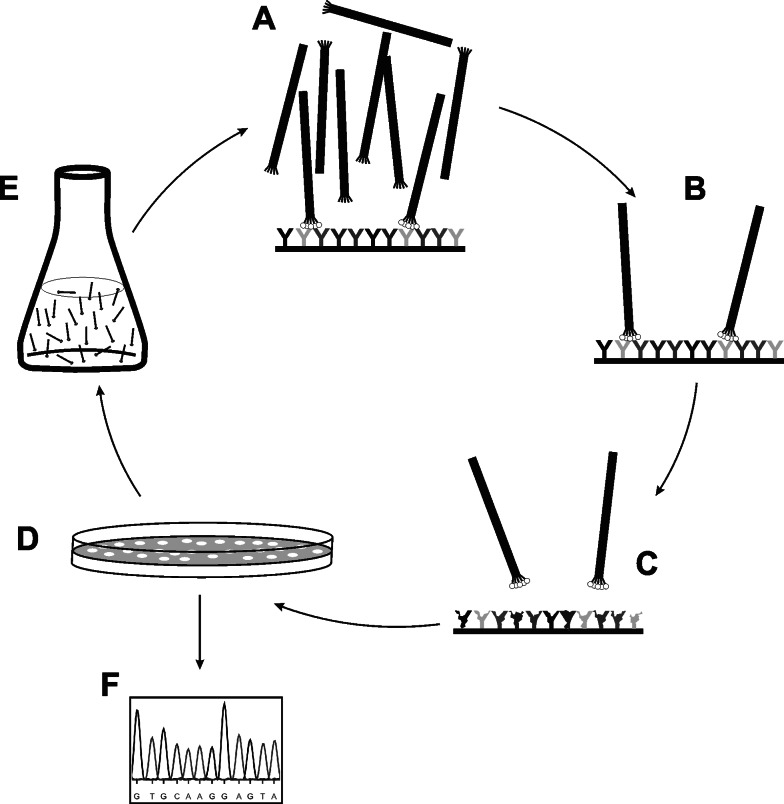
Basic principle of phage library screening. Library phage are incubated over an immobilized target (a). Non-binding phage are washed away (b) whereas target-binders are eluted (c). Eluted phage are plated on host bacteria (d), amplified in liquid growth medium (e) and subjected to further rounds of in vitro selection and in vivo amplification. Individual phage clones can be isolated from eluted subpopulation at any stage. Finally, primary structure of displayed ligand is easily identified via determination of insert’s nucleotide sequence (f). Usually, selected ligands (either on-phage or soluble synthetic/recombinant) are further analyzed to assess binding specificity and affinity (not shown)
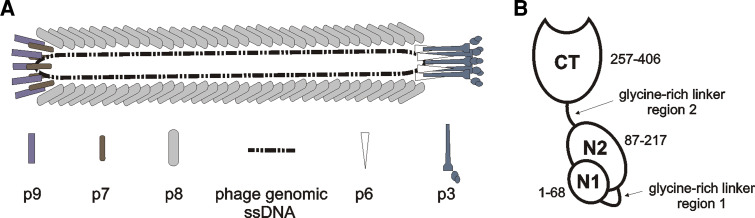
a Filamentous phage architecture (adapted from [21]). Filamentous phage are rod-like viruses with a circular ssDNA genome. The tube-like capsid is composed of several thousand copies of tightly packed major coat protein [gene VIII product (p8)], capped by five copies of p3 and p6 on one end and five copies of p7 and p9 on the opposite end. b Schematic representation of minor coat protein p3 structure (adapted from [21]). Two N-terminal domains (N2 and N1) are essential for phage infectivity. They are implicated in binding the F-pilus and TolA membrane protein of bacterial host, respectively, which act as phage receptors. The N-terminal part of p3 also renders infected cell immune to superinfection by another filamentous phage. C-terminal domain (CT) takes part in the final stages of virion assembly on inner bacterial membrane, capping the capsid end together with p6. Residue numbering is according to the mature p3
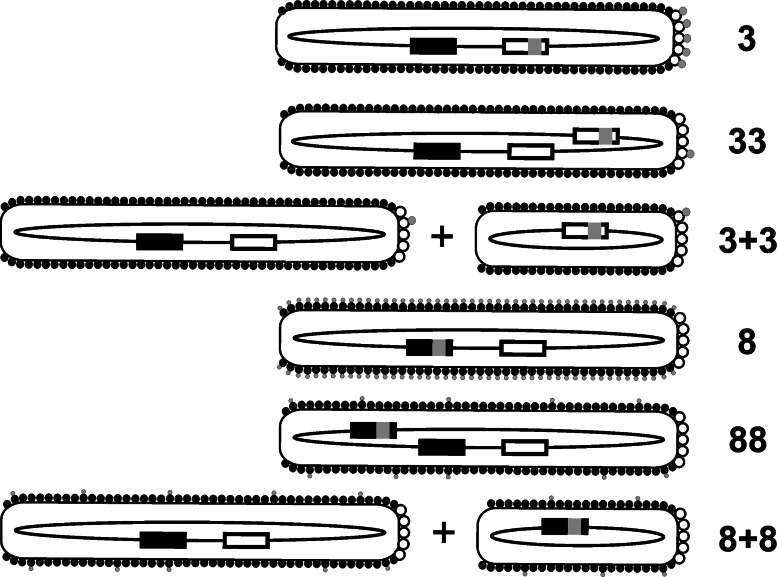
Classification of major phage display formats (adapted from [23]). White and black boxes represent wild-type gIII and gVIII, respectively. Foreign DNA inserts encoding displayed peptides (gray circles) are depicted by gray squares. White and black circles symbolize wild-type p3 and p8, respectively. In 3+3 and 8+8 format depictions, smaller phage represent phagemid virions, whereas larger ones depict helper phage. See text for details
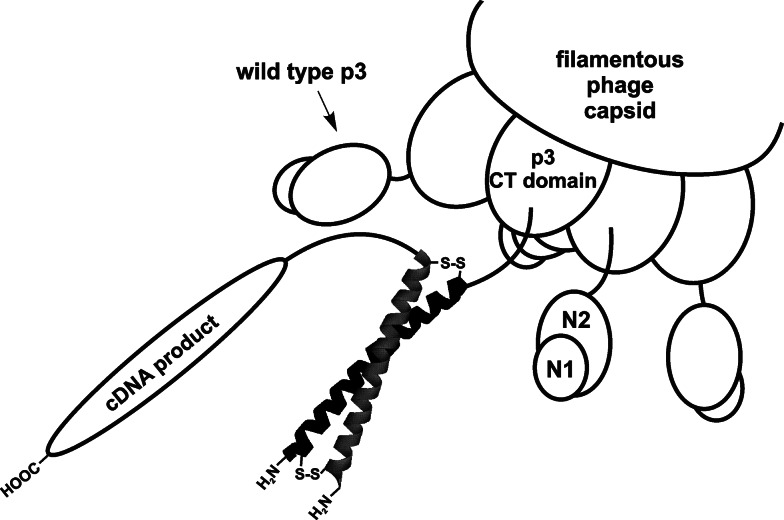
Depiction of p3-displayed cDNA products on a bicomponent Fos-Jun complex. Jun domain (black helix) is fused to minor capsid protein p3 while cDNA proteins are coupled to the C-terminus of Fos-domain (gray helix). Fos and Jun form a coiled coil complex (leucine zipper) reinforced with disulphide bonds
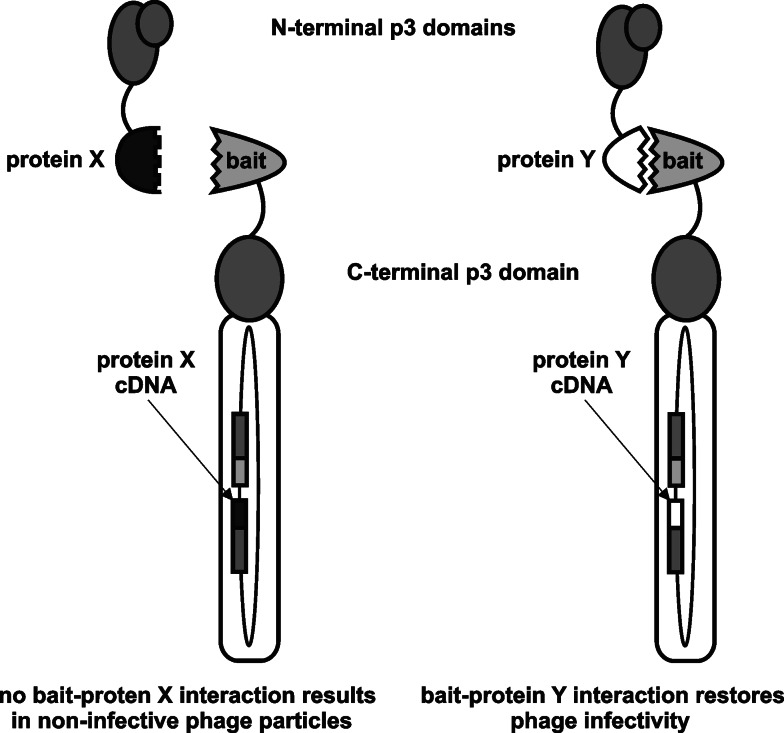
The principle of direct interaction rescue technique (adapted from [31]). Separate N- and C-terminal halves of p3 are rejoined through interaction of heterologous interacting protein domains, restoring phage infectivity
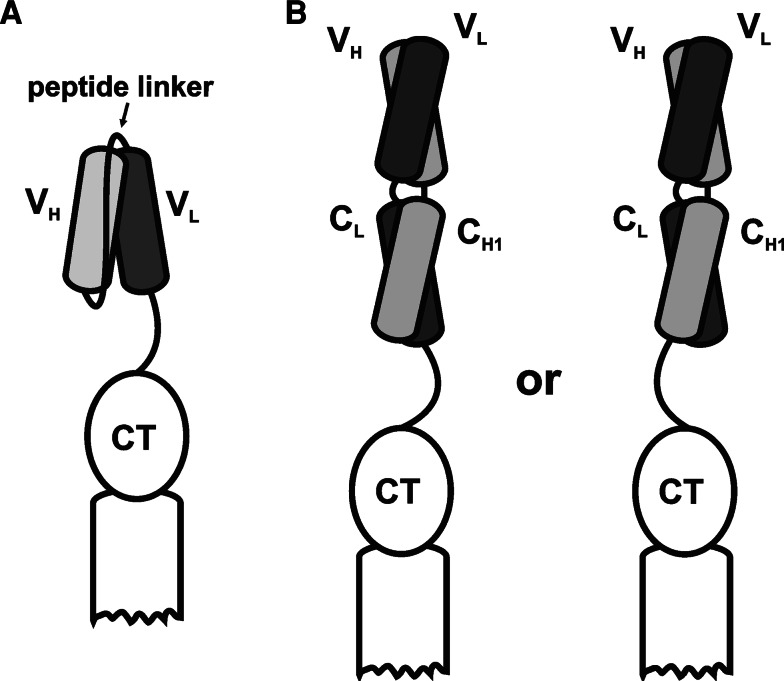
Schematic representation of antibody fragment types displayed as fusions to p3: a single-chain variable fragment (scFv); b antigen-binding fragment (Fab) with light chain-p3 fusion (left) or heavy chain-p3 fusion (right)
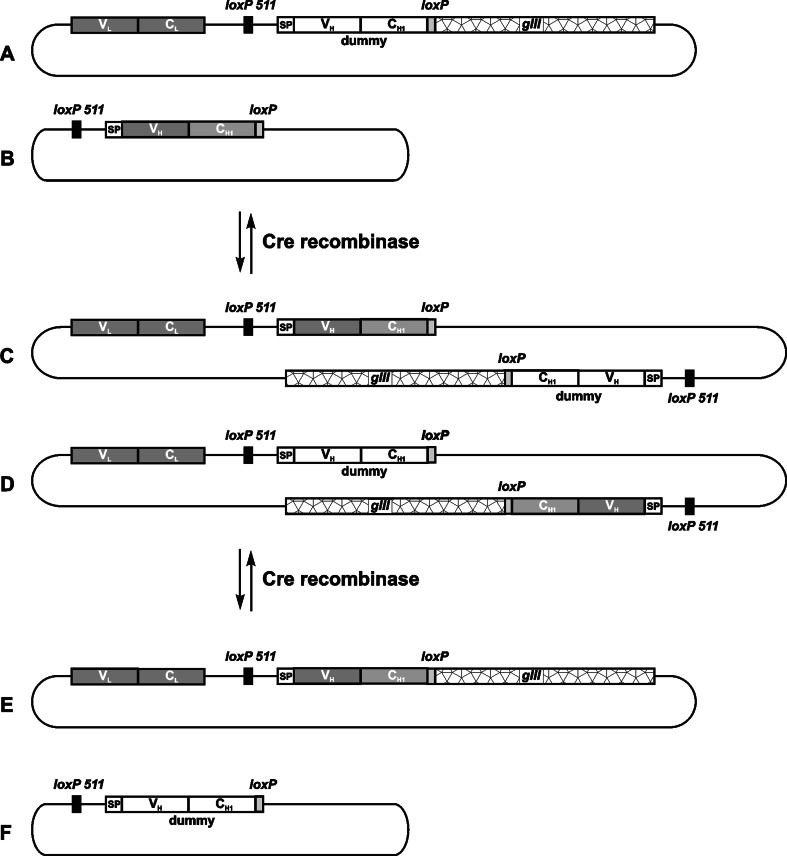
The principle of chain shuffling by in vivo recombination using the Cre-lox recombination system as reported by Waterhouse et al. [62]. Two replicons enter recombination: the acceptor fd phage vector (a), carrying the relevant light chain gene (VL–CL) and a dummy heavy chain gene (VH–CH1_dummy) fused to gIII, and the pUC19 donor plasmid (b), encoding the relevant heavy chain gene (VH-CH1). On both vectors, heavy chain genes are flanked by heterologous recombination sites (loxP and loxP 511). In the presence of Cre recombinase, chimaeric vectors are formed (c,d), which can undergo further recombination to yield either the original replicons (a,b) or two new vectors (e,f). Newly formed fd vector (e) now carries genetic information for a functional antigen-binding fragment
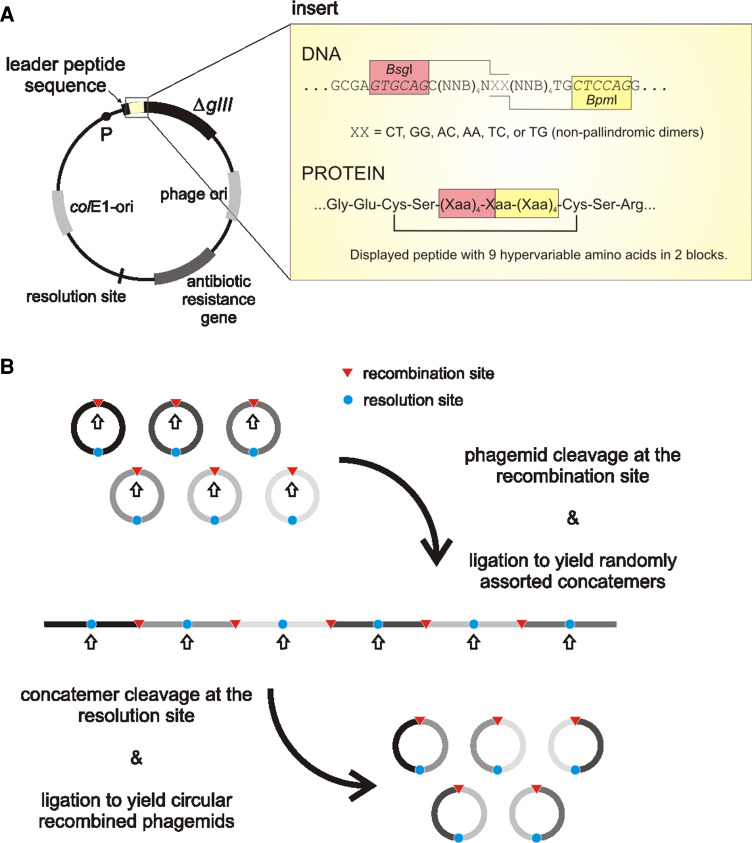
a General features of a cosmix-plexing phagemid vector. The insert of CPS1 library (a set of six sub-libraries) displaying cyclic (disulphide constrained) decapeptides is depicted as an example. The two hypervariable regions are separated by restriction sites BsgI/BpmI. In vitro recombination is induced by cleavage with either of the restrictases: in any case, a dinucleotide cohesive-end 3′-extension containing the sequence XX is produced. Limiting the sequence XX to one of the six shown, ensures that subsequent re-ligation only produces concatemers with the correct fragment orientation. P Promoter, ΔgIII gene encoding the C-terminal domain of p3, colE1-ori plasmid’s origin of replication, N any nucleotide (A, G, T, or C), B nucleotides C, G or T, Xaa any amino acid. b In vitro recombination procedure (adapted from [71])
References
-
- Pennazio S. The origin of phage virology. Riv Biol. 2006;99:103–129. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
