

Selective and potent furin inhibitors protect cells from anthrax without significant toxicity
- PMID: 20197107
- PMCID: PMC2862824
- DOI: 10.1016/j.biocel.2010.02.013
Selective and potent furin inhibitors protect cells from anthrax without significant toxicity
Abstract
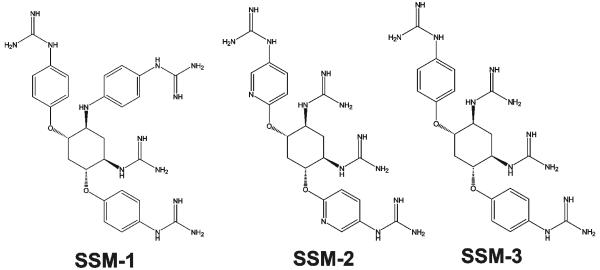
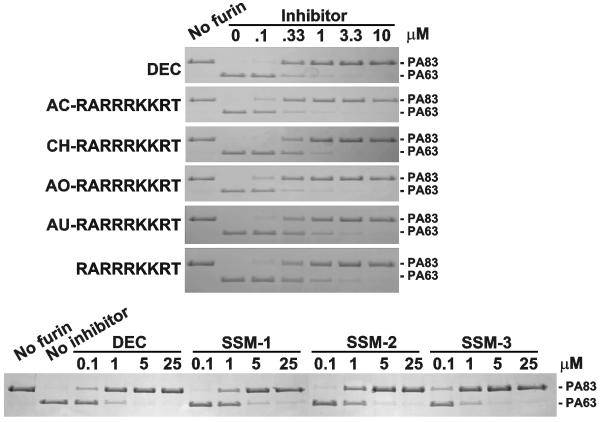
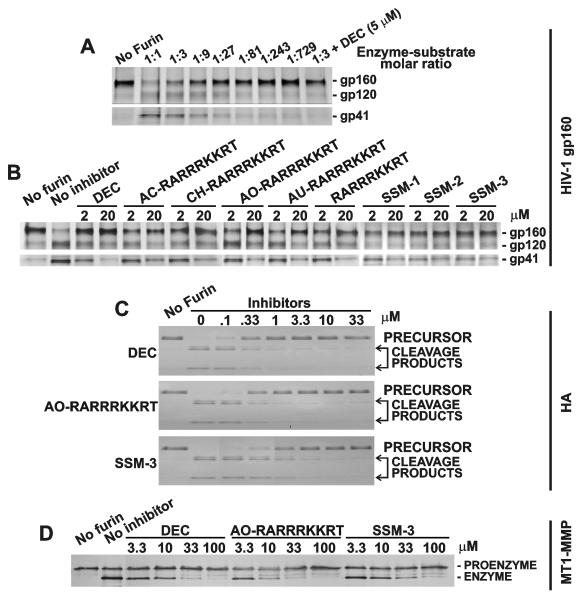
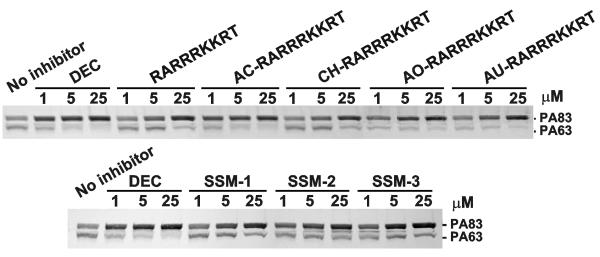
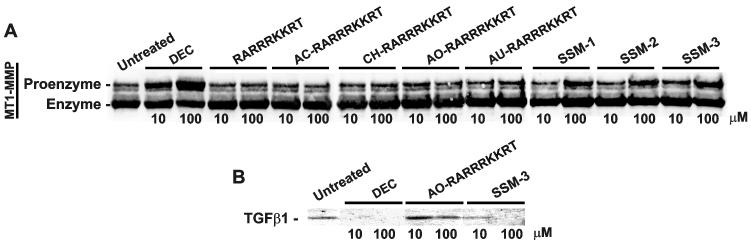
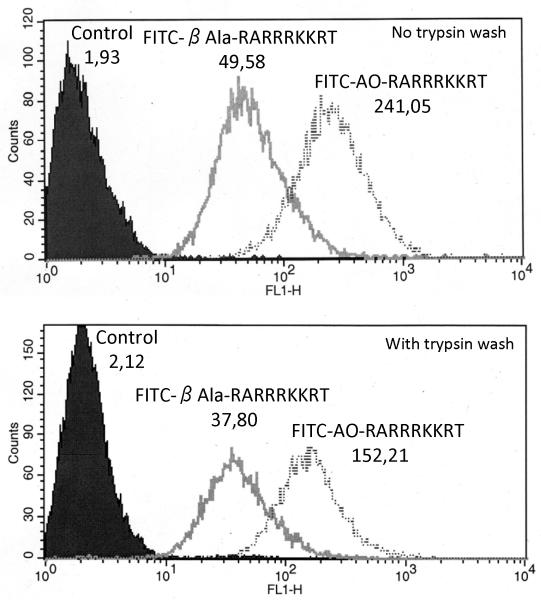
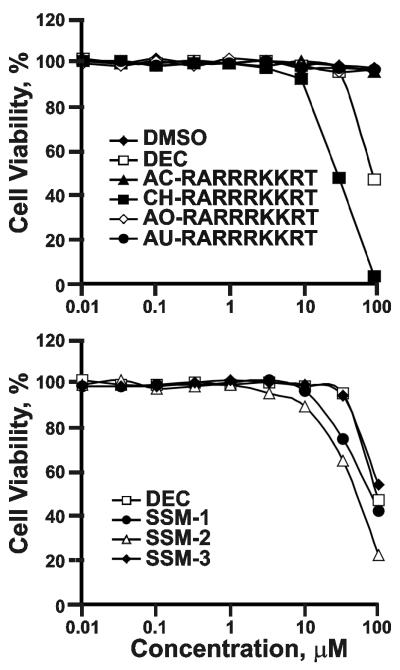
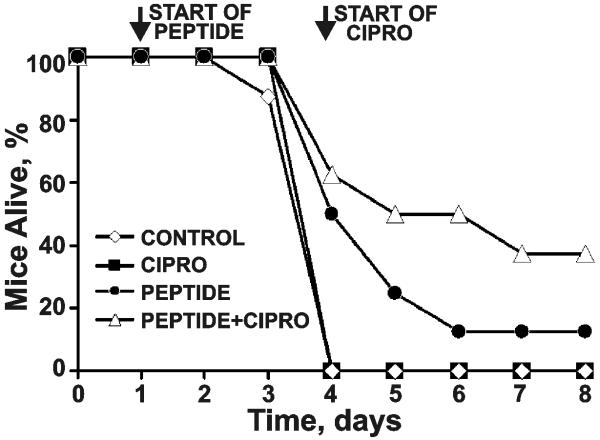
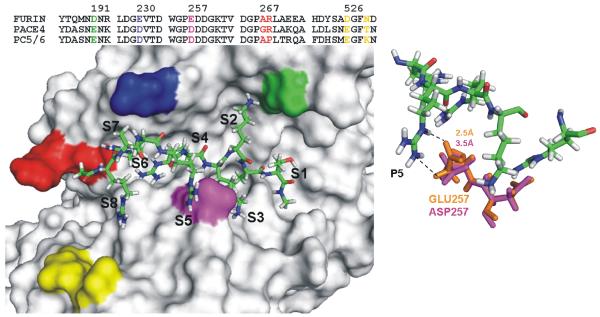
Furin and related proprotein convertases cleave the multibasic motifs R-X-R/K/X-R in the precursor proteins and, as a result, transform the latent proproteins into biologically active proteins and peptides. Furin is present both in the intracellular secretory pathway and at the cell surface. Intracellular furin processes its multiple normal cellular targets in the Golgi and secretory vesicle compartments while cell-surface furin appears to be essential only for the processing of certain pathogenic proteins and, importantly, anthrax. To design potent, safe and selective inhibitors of furin, we evaluated the potency and selectivity of the derivatized peptidic inhibitors modeled from the extended furin cleavage sequence of avian influenza A H5N1. We determined that the N- and C-terminal modifications of the original RARRRKKRT inhibitory scaffold produced selective and potent, nanomolar range, inhibitors of furin. These inhibitors did not interfere with the normal cellular function of furin because of the likely functional redundancy existing between furin and other proprotein convertases. These furin inhibitors, however, were highly potent in blocking the furin-dependent cell-surface processing of anthrax protective antigen-83 both in vitro and cell-based assays and in vivo. We conclude that the inhibitors we have designed have a promising potential as selective anthrax inhibitors, without affecting major cell functions.
Copyright 2010 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
A femtomol range FRET biosensor reports exceedingly low levels of cell surface furin: implications for the processing of anthrax protective antigen.PLoS One. 2010 Jun 24;5(6):e11305. doi: 10.1371/journal.pone.0011305. PLoS One. 2010. Retraction in: PLoS One. 2022 May 26;17(5):e0269294. doi: 10.1371/journal.pone.0269294. PMID: 20585585 Free PMC article. Retracted.
-
Anthrax toxin requires ZDHHC5-mediated palmitoylation of its surface-processing host enzymes.Proc Natl Acad Sci U S A. 2019 Jan 22;116(4):1279-1288. doi: 10.1073/pnas.1812588116. Epub 2019 Jan 4. Proc Natl Acad Sci U S A. 2019. PMID: 30610172 Free PMC article.
-
Targeting host cell furin proprotein convertases as a therapeutic strategy against bacterial toxins and viral pathogens.J Biol Chem. 2007 Jul 20;282(29):20847-53. doi: 10.1074/jbc.M703847200. Epub 2007 May 30. J Biol Chem. 2007. PMID: 17537721
-
New developments in vaccines, inhibitors of anthrax toxins, and antibiotic therapeutics for Bacillus anthracis.Curr Med Chem. 2011;18(33):5083-94. doi: 10.2174/092986711797636036. Curr Med Chem. 2011. PMID: 22050756 Free PMC article. Review.
-
New insights into the pathogenesis and treatment of anthrax toxin-induced shock.Expert Opin Biol Ther. 2007 Jun;7(6):843-54. doi: 10.1517/14712598.7.6.843. Expert Opin Biol Ther. 2007. PMID: 17555370 Review.
Cited by
-
Anthrax protective antigen is a calcium-dependent serine protease.Virulence. 2018;9(1):1085-1091. doi: 10.1080/21505594.2018.1486139. Virulence. 2018. PMID: 30052476 Free PMC article.
-
Domain flexibility modulates the heterogeneous assembly mechanism of anthrax toxin protective antigen.J Mol Biol. 2012 Jan 6;415(1):159-74. doi: 10.1016/j.jmb.2011.10.035. Epub 2011 Oct 31. J Mol Biol. 2012. PMID: 22063095 Free PMC article.
-
Does Covera-19 know 'when to hold 'em or 'when to fold 'em? A translational thought experiment.Transl Med Commun. 2021;6(1):12. doi: 10.1186/s41231-021-00090-5. Epub 2021 Jun 30. Transl Med Commun. 2021. PMID: 34226878 Free PMC article. Review.
-
GDF10 is a negative regulator of vascular calcification.J Biol Chem. 2024 Nov;300(11):107805. doi: 10.1016/j.jbc.2024.107805. Epub 2024 Sep 21. J Biol Chem. 2024. PMID: 39307303 Free PMC article.
-
AB toxins: a paradigm switch from deadly to desirable.Toxins (Basel). 2010 Jul;2(7):1612-45. doi: 10.3390/toxins2071612. Epub 2010 Jun 25. Toxins (Basel). 2010. PMID: 22069653 Free PMC article. Review.
References
-
- Anderson ED, Thomas L, Hayflick JS, Thomas G. Inhibition of HIV-1 gp160-dependent membrane fusion by a furin-directed alpha 1-antitrypsin variant. J Biol Chem. 1993;268:24887–91. - PubMed
-
- Basak A, Zhong M, Munzer JS, Chretien M, Seidah NG. Implication of the proprotein convertases furin, PC5 and PC7 in the cleavage of surface glycoproteins of Hong Kong, Ebola and respiratory syncytial viruses: a comparative analysis with fluorogenic peptides. Biochem J. 2001;353:537–45. - PMC - PubMed
-
- Bassi DE, Fu J, de Cicco R Lopez, Klein-Szanto AJ. Proprotein convertases: “master switches” in the regulation of tumor growth and progression. Mol Carcinog. 2005;44:151–61. - PubMed
-
- Benjannet S, Savaria D, Laslop A, Munzer JS, Chretien M, Marcinkiewicz M, Seidah NG. Alpha1-antitrypsin Portland inhibits processing of precursors mediated by proprotein convertases primarily within the constitutive secretory pathway. J Biol Chem. 1997;272:26210–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 AI059572/AI/NIAID NIH HHS/United States
- R01 CA077470/CA/NCI NIH HHS/United States
- U01 AI078048/AI/NIAID NIH HHS/United States
- R01 CA083017/CA/NCI NIH HHS/United States
- U01 AI056385/AI/NIAID NIH HHS/United States
- CAPMC/ CIHR/Canada
- CA83017/CA/NCI NIH HHS/United States
- RR020843/RR/NCRR NIH HHS/United States
- AI055789/AI/NIAID NIH HHS/United States
- CA77470/CA/NCI NIH HHS/United States
- AI078048/AI/NIAID NIH HHS/United States
- U54 RR020843/RR/NCRR NIH HHS/United States
- P01 AI055789/AI/NIAID NIH HHS/United States
- AI059572/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
