

Review
doi: 10.1128/MMBR.00040-09.
Getting in the loop: regulation of development in Caulobacter crescentus
Affiliations
- PMID: 20197497
- PMCID: PMC2832345
- DOI: 10.1128/MMBR.00040-09
Item in Clipboard
Review
Getting in the loop: regulation of development in Caulobacter crescentus
Microbiol Mol Biol Rev.
2010 Mar.
Abstract
Caulobacter crescentus is an aquatic Gram-negative alphaproteobacterium that undergoes multiple changes in cell shape, organelle production, subcellular distribution of proteins, and intracellular signaling throughout its life cycle. Over 40 years of research has been dedicated to this organism and its developmental life cycles. Here we review a portion of many developmental processes, with particular emphasis on how multiple processes are integrated and coordinated both spatially and temporally. While much has been discovered about Caulobacter crescentus development, areas of potential future research are also highlighted.
Figures
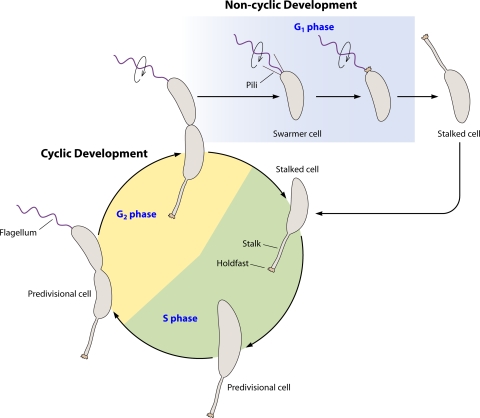
Life cycle of Caulobacter crescentus. The cyclic developmental program begins with a stalked cell with an adhesive holdfast at the tip of the stalk. The stalked cell enters S phase, a cell state where it is competent for DNA replication. As the cell grows and replicates its DNA, it becomes a predivisional cell. During this time the cell becomes incompetent for DNA replication, entering the G2 phase. In the late predivisional stage, a flagellum is formed at the swarmer cell pole. After compartmentalization, flagellar rotation is activated (circular arrow) and pili are extruded. Cell separation leads to two different cell types. One cell is a stalked cell which reenters the cyclic developmental program and S phase, completing the circle. The other cell is a swarmer cell. The swarmer cell cannot replicate its chromosome yet is distinct from the predivisional cell and therefore is in a separate phase, referred to as G1. The holdfast is formed predominantly during the swarmer cell stage. Later the swarmer cell differentiates into a stalked cell. This differentiation comprises the noncyclic developmental program.
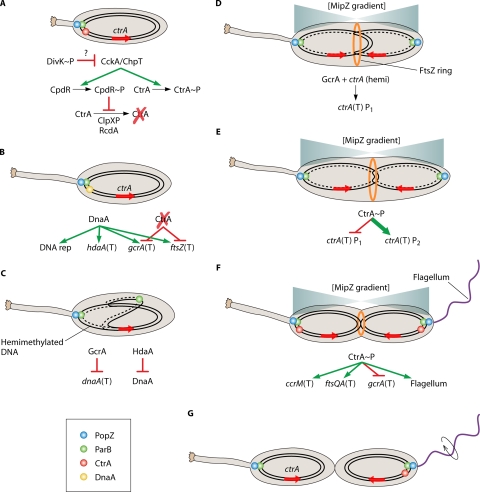
Integral cell cycle components of the cyclic developmental program. (A) The initiation of the cyclic developmental program occurs during swarmer cell differentiation or in stalked cells that are the product of cell division, and it requires deactivation of CtrA. In the stalked cell that is the product of cell division, CtrA is deactivated after compartmentalization. In the stalked cell that is the product of swarmer cell differentiation (as indicated by a shortened stalk), activated CtrA (phosphorylated and not proteolytically degraded) is bound to and silences the origin of replication and must be deactivated (see “Modulation of CtrA Activity” in the text). DivK∼P leads to inhibition of CckA-mediated activation of CtrA by an unknown mechanism. Phosphoryl transfer from CckA to ChpT to CtrA ceases, preventing CtrA phosphorylation. Phosphoryl transfer from CckA to ChpT to CpdR also ceases, leading to decreased CpdR∼P levels and relieving inhibition of CtrA proteolysis. CtrA becomes deactivated (dephosphorylated and proteolytically degraded). In stalked cells originating from swarmer cell differentiation or cell division, the ParB-parS complex is bound to PopZ multimers at the stalked cell pole. (B) DnaA is synthesized and binds to the origin of replication vacated by CtrA, initiating DNA replication. DnaA also positively regulates transcription (T) of hdaA, gcrA, and ftsZ. The absence of activated CtrA allows transcription of gcrA as well as ftsZ. (C) One of the new ParB-parS complexes begins migration across the cell to the swarmer pole. DNA replication continues, leaving the replicated DNA in the hemimethylated state. GcrA represses transcription of dnaA, while HdaA inactivates existing DnaA, preventing additional initiation of DNA replication. (D) Segregated ParB-parS complexes allow establishment of MipZ gradients, with the lowest concentration at roughly midcell, determining the position of the FtsZ ring (orange ring). DNA replication past ctrA (red arrow) leaves the duplicated genes in the hemimethylated (hemi) state. Hemimethylation in combination with GcrA lead to activation of the weak ctrA P1 promoter. (E) The small amount of CtrA produced and activated represses the P1 promoter and activates the strong ctrA P2 promoter, leading a burst of CtrA synthesis (bold). (F) The burst of CtrA synthesis has multiple effects on the cell, including silencing the origin of replication, initiating CcrM synthesis which methylates the chromosomes, initiating FtsQA synthesis which allows cytokinesis to begin, blocking gcrA transcription, initiating flagellum biosynthesis, and other effects as well. (G) Once cytokinesis has completed, flagellum rotation is activated (circular arrow), and the two new cells are ready to separate. CtrA is deactivated in the stalked cell, allowing reinitiation of the cyclic developmental program. The mechanism for cell type-specific CtrA inactivation is described in Fig. 3.
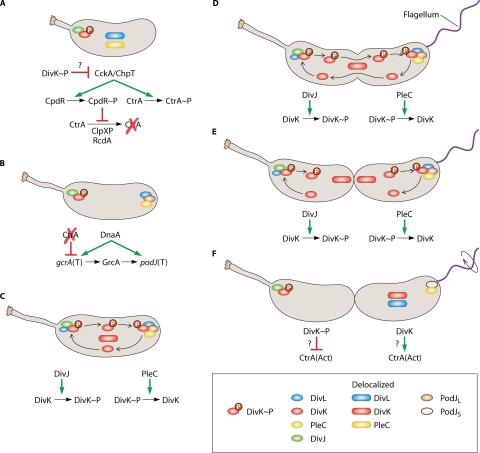
Cytokinesis-sensing mechanism. (A) As shown in Fig. 2, the developmental cycle represented begins with either a differentiating swarmer cell (as indicated by the shortened stalk) or a stalked cell immediately after cell division. The cyclic developmental cycle begins with DivJ and DivK∼P localized to the stalked pole. DivL and PleC are delocalized in the inner membrane. As described for Fig. 2, DivK∼P leads to deactivation of CtrA. (B) DnaA and the absence of activated CtrA lead to gcrA transcription (T). DnaA and GcrA lead to production of PodJ. PodJ localizes to the nascent swarmer pole and serves as a localization factor for PleC. DivL also becomes localized to this pole. (C) DivJ and PleC are located at opposite poles. DivK is phosphorylated by DivJ at the stalked pole and localizes there. DivL is found predominantly at the swarmer pole in the predivisional cell but is sometimes detected as a dimmer focus as the stalked pole, indicated by a smaller circle (see “DivL: a Wrench in the Works” in the text). This inconsistent DivL detection at the stalked pole could indicate a smaller protein pool at this location or a transient localization state. In either situation, DivL could serve as a DivK∼P localization factor at this pole. DivK∼P also diffuses to the swarmer pole (arrow trails) and forms a focus, again potentially through DivL. PleC dephosphorylates DivK∼P at the swarmer pole, causing it to delocalize. Delocalized DivK diffuses (arrow trails) and becomes rephosphorylated by DivJ at the stalked pole. (D) DivK phosphorylation/dephosphorylation cycling continues as the cell cycle progresses. (E) Immediately after cytokinesis completes, compartmentalization isolates DivJ and PleC enzymatic activities from each other. (F) As a consequence of compartmentalization, isolated DivJ activity leads to DivK phosphorylation in the stalked cell compartment, which then polarly localizes. Isolated PleC activity leads to DivK dephosphorylation and subsequently delocalization. The phosphorylation state of DivK affects activation/inactivation of CtrA in each cell compartment. CtrA activation/inactivation is accomplished by modulation of the CckA pathway, and though it is unknown exactly how DivK(∼P) interacts with this pathway, DivK∼P favors CtrA inactivation (see panel A). Conversely, DivK favors CtrA activation, perhaps simply by not favoring its inactivation, although other mechanisms have not been ruled out. As a result, CtrA is deactivated in the stalked cell compartment, while CtrA is activated in the swarmer cell compartment. PodJ is proteolytically processed to the short form, though PleC remains localized, and DivL and DivK become delocalized. Flagellar rotation is activated (circular arrow).
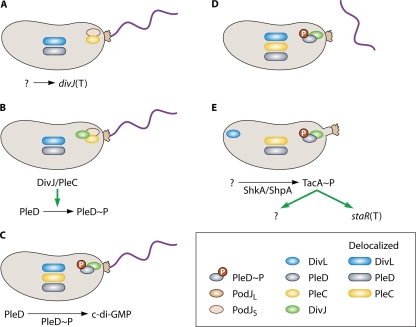
Swarmer cell differentiation. (A) A mature swarmer cell is indicated by the presence of holdfast at the flagellar pole and the absence of pili. PodJS and PleC are localized to the flagellar pole. PleD is in the nonphosphorylated state and therefore delocalized, as is DivL. An unknown signal leads to DivJ synthesis. (B) DivJ is synthesized and localizes to the flagellar pole. For a brief period of time, PodJs, PleC, and DivJ all inhabit the same pole. DivJ and/or PleC phosphorylate PleD (see “Swarmer → Stalked Cell Transition” in the text), causing a subpopulation to localize to the developing pole. (C) PodJS is degraded by an unknown mechanism, and PleC becomes delocalized. PleD∼P catalyzes the formation of c-di-GMP. (D) Production of c-di-GMP leads to morphological changes in the cell through unknown mechanisms. The flagellum is ejected. (E) Stalk synthesis is initiated, which requires phosphorylation of the sigma 54 activator TacA through the ShkA/ShpA phosphorelay. The signal that leads to ShkA/ShpA/TacA activation is unknown. TacA activation leads to synthesis of the stalk length determinant StaR, as well as other, unidentified targets. Other factors are likely involved in stalk synthesis. It should be noted that the ShkA/ShpA/TacA pathway is present and active in the late predivisional and swarmer cell stages. The mechanism for controlling the timing of stalk synthesis is not understood. The stalk is extended with the holdfast at the tip. DivL localizes to the stalk-distal pole.
References
-
- Aaron, M., G. Charbon, H. Lam, H. Schwarz, W. Vollmer, and C. Jacobs-Wagner. 2007. The tubulin homologue FtsZ contributes to cell elongation by guiding cell wall precursor synthesis in Caulobacter crescentus. Mol. Microbiol. 64:938-952. - PubMed
-
- Aldridge, P., and K. T. Hughes. 2002. Regulation of flagellar assembly. Curr. Opin. Microbiol. 5:160-165. - PubMed
-
- Aldridge, P., and U. Jenal. 1999. Cell cycle-dependent degradation of a flagellar motor component requires a novel-type response regulator. Mol. Microbiol. 32:379-391. - PubMed
-
- Aldridge, P., R. Paul, P. Goymer, P. Rainey, and U. Jenal. 2003. Role of the GGDEF regulator PleD in polar development of Caulobacter crescentus. Mol. Microbiol. 47:1695-1708. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
