

Molecular mechanism of flop selectivity and subsite recognition for an AMPA receptor allosteric modulator: structures of GluA2 and GluA3 in complexes with PEPA
- PMID: 20199107
- PMCID: PMC3072023
- DOI: 10.1021/bi1000678
Molecular mechanism of flop selectivity and subsite recognition for an AMPA receptor allosteric modulator: structures of GluA2 and GluA3 in complexes with PEPA
Abstract
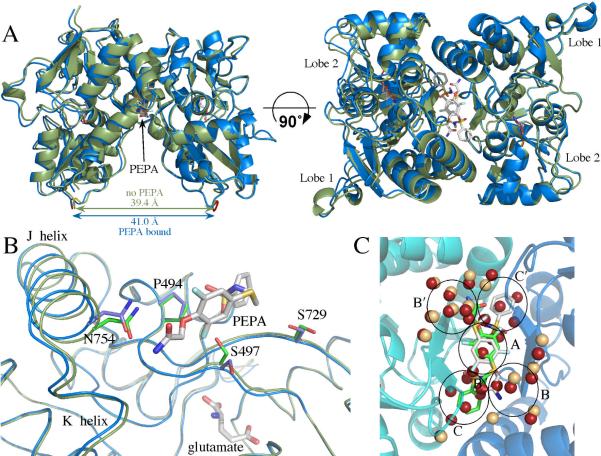
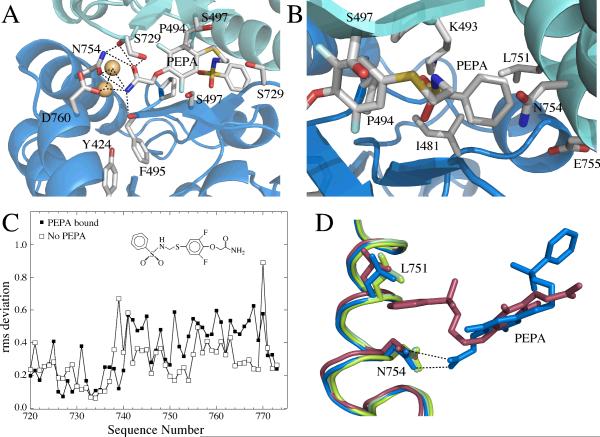
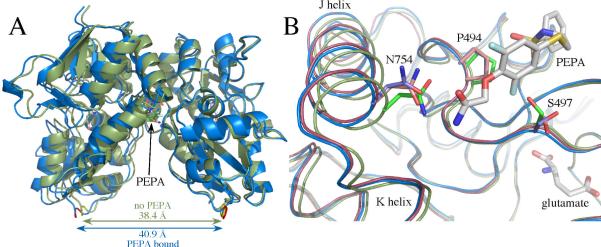
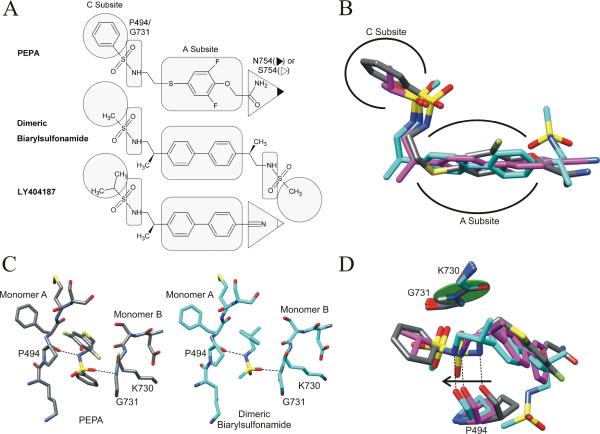
Glutamate receptors are important potential drug targets for cognitive enhancement and the treatment of schizophrenia in part because they are the most prevalent excitatory neurotransmitter receptors in the vertebrate central nervous system. One approach to the application of therapeutic agents to the AMPA subtype of glutamate receptors is the use of allosteric modulators, which promote dimerization by binding to a dimer interface thereby reducing the degree of desensitization and deactivation. AMPA receptors exist in two alternatively spliced variants (flip and flop) that differ in desensitization and receptor activation profiles. Most of the structural information about modulators of the AMPA receptor targets the flip subtype. We report here the crystal structure of the flop-selective allosteric modulator, PEPA, bound to the binding domains of the GluA2 and GluA3 flop isoforms of AMPA receptors. Specific hydrogen bonding patterns can explain the preference for the flop isoform. This includes a bidentate hydrogen bonding pattern between PEPA and N754 of the flop isoforms of GluA2 and GluA3 (the corresponding position in the flip isoform is S754). Comparison with other allosteric modulators provides a framework for the development of new allosteric modulators with preferences for either the flip or flop isoforms. In addition to interactions with N/S754, specific interactions of the sulfonamide with conserved residues in the binding site are characteristics of a number of allosteric modulators. These, in combination with variable interactions with five subsites on the binding surface, lead to different stoichiometries, orientations within the binding pockets, and functional outcomes.
Figures
References
-
- Christopoulos A. Allosteric binding sites on cell-surface receptors: novel targets for drug discovery. Nat Rev Drug Discov. 2002;1:198–210. - PubMed
-
- Changeux JP, Taly A. Nicotinic receptors, allosteric proteins and medicine. Trends Mol Med. 2008;14:93–102. - PubMed
-
- Dingledine R, Borges K, Bowie D, Traynelis S. The glutamate receptor ion channels. Pharmacol. Rev. 1999;51:7–61. - PubMed
-
- Oswald RE, Ahmed A, Fenwick MK, Loh AP. Structure of glutamate receptors. Current drug targets. 2007;8:573–582. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
