

Auxin metabolism and function in the multicellular brown alga Ectocarpus siliculosus
- PMID: 20200071
- PMCID: PMC2862433
- DOI: 10.1104/pp.109.149708
Auxin metabolism and function in the multicellular brown alga Ectocarpus siliculosus
Abstract
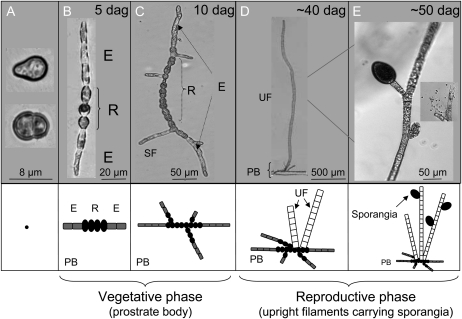
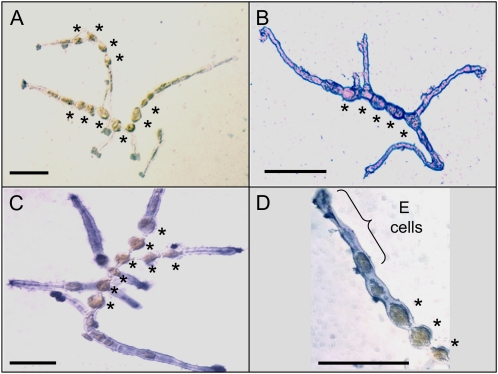
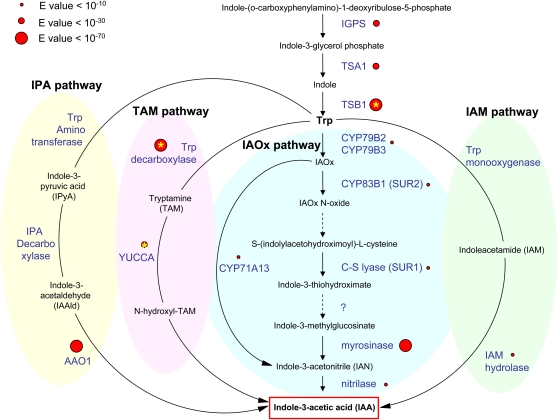
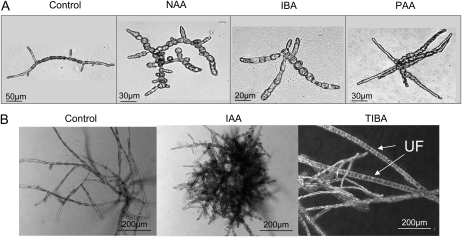
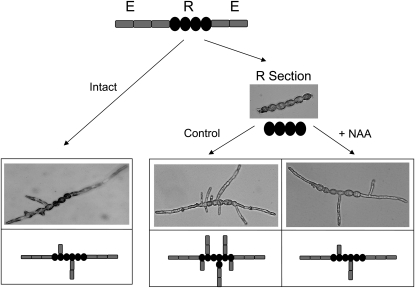
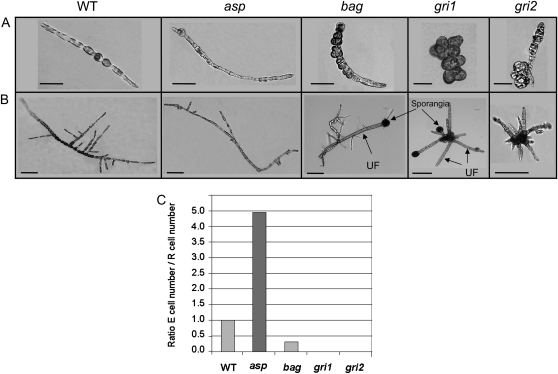
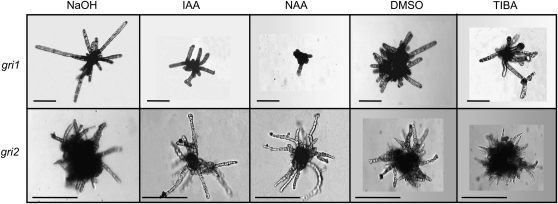
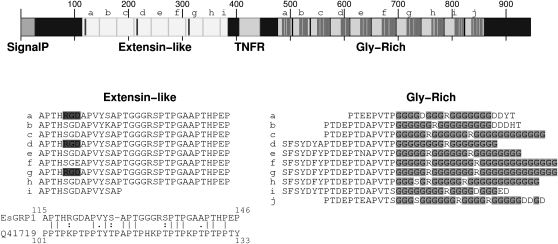
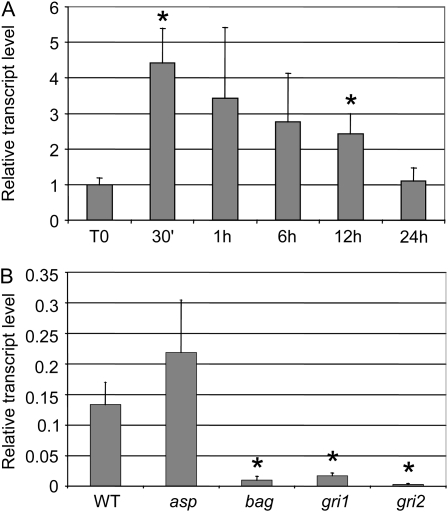
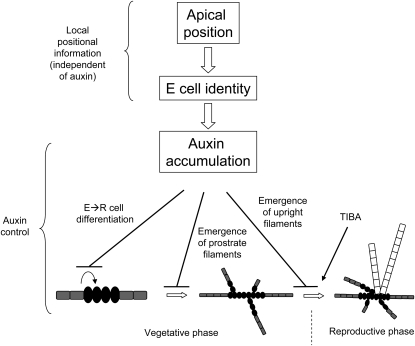
Ectocarpus siliculosus is a small brown alga that has recently been developed as a genetic model. Its thallus is filamentous, initially organized as a main primary filament composed of elongated cells and round cells, from which branches differentiate. Modeling of its early development suggests the involvement of very local positional information mediated by cell-cell recognition. However, this model also indicates that an additional mechanism is required to ensure proper organization of the branching pattern. In this paper, we show that auxin indole-3-acetic acid (IAA) is detectable in mature E. siliculosus organisms and that it is present mainly at the apices of the filaments in the early stages of development. An in silico survey of auxin biosynthesis, conjugation, response, and transport genes showed that mainly IAA biosynthesis genes from land plants have homologs in the E. siliculosus genome. In addition, application of exogenous auxins and 2,3,5-triiodobenzoic acid had different effects depending on the developmental stage of the organism, and we propose a model in which auxin is involved in the negative control of progression in the developmental program. Furthermore, we identified an auxin-inducible gene called EsGRP1 from a small-scale microarray experiment and showed that its expression in a series of morphogenetic mutants was positively correlated with both their elongated-to-round cell ratio and their progression in the developmental program. Altogether, these data suggest that IAA is used by the brown alga Ectocarpus to relay cell-cell positional information and induces a signaling pathway different from that known in land plants.
Figures

Similar articles
-
Auxin Function in the Brown Alga Dictyota dichotoma.Plant Physiol. 2019 Jan;179(1):280-299. doi: 10.1104/pp.18.01041. Epub 2018 Nov 12. Plant Physiol. 2019. PMID: 30420566 Free PMC article.
-
Space-time decoupling in the branching process in the mutant étoile of the filamentous brown alga Ectocarpus siliculosus.Plant Signal Behav. 2011 Dec;6(12):1889-92. doi: 10.4161/psb.6.12.18054. Plant Signal Behav. 2011. PMID: 22095146 Free PMC article.
-
ETOILE regulates developmental patterning in the filamentous brown alga Ectocarpus siliculosus.Plant Cell. 2011 Apr;23(4):1666-78. doi: 10.1105/tpc.110.081919. Epub 2011 Apr 8. Plant Cell. 2011. PMID: 21478443 Free PMC article.
-
Development and physiology of the brown alga Ectocarpus siliculosus: two centuries of research.New Phytol. 2008;177(2):319-332. doi: 10.1111/j.1469-8137.2007.02304.x. New Phytol. 2008. PMID: 18181960 Review.
-
[A Picasso among brown algae: the arduous conquest of symmetry by Ectocarpus].J Soc Biol. 2007;201(3):267-80. doi: 10.1051/jbio:2007038. J Soc Biol. 2007. PMID: 18157079 Review. French.
Cited by
-
The evolution of the plant genome-to-morphology auxin circuit.Theory Biosci. 2016 Sep;135(3):175-86. doi: 10.1007/s12064-016-0231-0. Epub 2016 Jun 22. Theory Biosci. 2016. PMID: 27333773
-
Understanding "green" multicellularity: do seaweeds hold the key?Front Plant Sci. 2015 Jan 20;5:737. doi: 10.3389/fpls.2014.00737. eCollection 2014. Front Plant Sci. 2015. PMID: 25653653 Free PMC article. No abstract available.
-
Auxin Function in the Brown Alga Dictyota dichotoma.Plant Physiol. 2019 Jan;179(1):280-299. doi: 10.1104/pp.18.01041. Epub 2018 Nov 12. Plant Physiol. 2019. PMID: 30420566 Free PMC article.
-
The birth of a giant: evolutionary insights into the origin of auxin responses in plants.EMBO J. 2023 Mar 15;42(6):e113018. doi: 10.15252/embj.2022113018. Epub 2023 Feb 14. EMBO J. 2023. PMID: 36786017 Free PMC article. Review.
-
Towards an understanding of spiral patterning in the Sargassum muticum shoot apex.Sci Rep. 2017 Oct 24;7(1):13887. doi: 10.1038/s41598-017-13767-5. Sci Rep. 2017. PMID: 29066850 Free PMC article.
References
-
- Abad MJ, Bedoya LM, Bermejo P. (2008) Natural marine anti-inflammatory products. Mini Rev Med Chem 8: 740–754 - PubMed
-
- Armbrust EV, Berges JA, Bowler C, Green BR, Martinez D, Putnam NH, Zhou S, Allen AE, Apt KE, Bechner M, et al. (2004) The genome of the diatom Thalassiosira pseudonana: ecology, evolution, and metabolism. Science 306: 79–86 - PubMed
-
- Baldauf SL. (2008) An overview of the phylogeny and diversity of eukaryotes. J Syst Evol 46: 263–273
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
