

Stromal hyaluronan interaction with epithelial CD44 variants promotes prostate cancer invasiveness by augmenting expression and function of hepatocyte growth factor and androgen receptor
- PMID: 20200161
- PMCID: PMC2888393
- DOI: 10.1074/jbc.M110.104273
Stromal hyaluronan interaction with epithelial CD44 variants promotes prostate cancer invasiveness by augmenting expression and function of hepatocyte growth factor and androgen receptor
Abstract
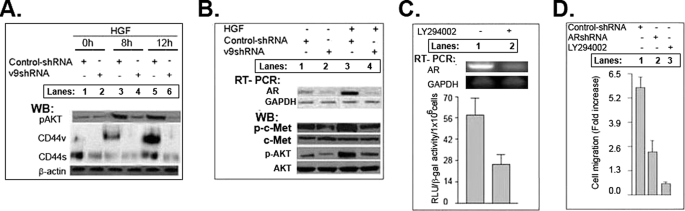
The main aim of our study is to determine the significance of the stromal microenvironment in the malignant behavior of prostate cancer. The stroma-derived growth factors/cytokines and hyaluronan act in autocrine/paracrine ways with their receptors, including receptor-tyrosine kinases and CD44 variants (CD44v), to potentiate and support tumor epithelial cell survival. Overexpression of hyaluronan, CD44v9 variants, and stroma-derived growth factors/cytokines are specific features in many cancers, including prostate cancer. Androgen/androgen receptor interaction has a critical role in regulating prostate cancer growth. Our previous study showed that 1) that increased synthesis of hyaluronan in normal epithelial cells promotes expression of CD44 variants; 2) hyaluronan interaction with CD44v6-v9 promotes activation of receptor-tyrosine kinase, which stimulates phosphatidylinositol 3-kinase-induced cell survival pathways; and 3) CD44v6/short hairpin RNA reduces colon tumor growth in vivo (Misra, S., Hascall, V. C., De Giovanni, C., Markwald, R. R., and Ghatak, S. (2009) J. Biol. Chem. 284, 12432-12446). Our results now show that hepatocyte growth factor synthesized by myofibroblasts associated with prostate cancer cells induces activation of HGF-receptor/cMet and stimulates hyaluronan/CD44v9 signaling. This, in turn, stabilizes the androgen receptor functions in prostate cancer cells. The stroma-derived HGF induces a lipid raft-associated signaling complex that contains CD44v9, cMet/phosphatidylinositol 3-kinase, HSP90 and androgen receptor. CD44v9/short hairpin RNA reverses the assembly of these components in the complex and inhibits androgen receptor function. Our results provide new insight into the hyaluronan/CD44v9-regulated androgen receptor function and the consequent malignant activities in prostate cancer cells. The present study describes a physiologically relevant in vitro model for studying the molecular mechanisms by which stroma-derived HGF and hyaluronan influence androgen receptor and CD44 functions in the secretory epithelia during prostate carcinogenesis.
Figures








Similar articles
-
Normal and malignant prostate epithelial cells differ in their response to hepatocyte growth factor/scatter factor.Am J Pathol. 2001 Aug;159(2):579-90. doi: 10.1016/S0002-9440(10)61729-4. Am J Pathol. 2001. PMID: 11485916 Free PMC article.
-
MET Signaling Mediates Intestinal Crypt-Villus Development, Regeneration, and Adenoma Formation and Is Promoted by Stem Cell CD44 Isoforms.Gastroenterology. 2017 Oct;153(4):1040-1053.e4. doi: 10.1053/j.gastro.2017.07.008. Epub 2017 Jul 14. Gastroenterology. 2017. PMID: 28716720
-
Hyaluronan-CD44 interaction promotes c-Src-mediated twist signaling, microRNA-10b expression, and RhoA/RhoC up-regulation, leading to Rho-kinase-associated cytoskeleton activation and breast tumor cell invasion.J Biol Chem. 2010 Nov 19;285(47):36721-35. doi: 10.1074/jbc.M110.162305. Epub 2010 Sep 15. J Biol Chem. 2010. PMID: 20843787 Free PMC article.
-
Hepatocyte growth factor/MET and CD44 in colorectal cancer: partners in tumorigenesis and therapy resistance.Biochim Biophys Acta Rev Cancer. 2020 Dec;1874(2):188437. doi: 10.1016/j.bbcan.2020.188437. Epub 2020 Sep 23. Biochim Biophys Acta Rev Cancer. 2020. PMID: 32976979 Review.
-
The CD147-HYALURONAN Axis in Cancer.Anat Rec (Hoboken). 2020 Jun;303(6):1573-1583. doi: 10.1002/ar.24147. Epub 2019 May 24. Anat Rec (Hoboken). 2020. PMID: 31090215 Review.
Cited by
-
CD44 integrates signaling in normal stem cell, cancer stem cell and (pre)metastatic niches.Exp Biol Med (Maywood). 2013 Mar;238(3):324-38. doi: 10.1177/1535370213480714. Exp Biol Med (Maywood). 2013. PMID: 23598979 Free PMC article. Review.
-
Investigation of androgen receptor-dependent alternative splicing has identified a unique subtype of lethal prostate cancer.Asian J Androl. 2023 May-Jun;25(3):296-308. doi: 10.4103/aja202263. Asian J Androl. 2023. PMID: 36259569 Free PMC article.
-
Understanding Hyaluronan Receptor (CD44) Interaction, HA-CD44 Activated Potential Targets in Cancer Therapeutics.Adv Pharm Bull. 2021 May;11(3):426-438. doi: 10.34172/apb.2021.050. Epub 2020 Jul 15. Adv Pharm Bull. 2021. PMID: 34513617 Free PMC article. Review.
-
CD44ICD promotes breast cancer stemness via PFKFB4-mediated glucose metabolism.Theranostics. 2018 Nov 29;8(22):6248-6262. doi: 10.7150/thno.28721. eCollection 2018. Theranostics. 2018. PMID: 30613295 Free PMC article.
-
Castrated autoimmune glomerulonephritis mouse model shows attenuated glomerular sclerosis with altered parietal epithelial cell phenotype.Exp Biol Med (Maywood). 2021 Jun;246(11):1318-1329. doi: 10.1177/1535370221996010. Epub 2021 Feb 27. Exp Biol Med (Maywood). 2021. PMID: 33641441 Free PMC article.
References
-
- Jemal A., Siegel R., Ward E., Hao Y., Xu J., Murray T., Thun M. J. (2008) CA Cancer J. Clin. 58, 71–96 - PubMed
-
- Grossmann M. E., Huang H., Tindall D. J. (2001) J. Natl. Cancer Inst. 93, 1687–1697 - PubMed
-
- Aaltomaa S., Lipponen P., Tammi R., Tammi M., Viitanen J., Kankkunen J. P., Kosma V. M. (2002) Urol. Int. 69, 266–272 - PubMed
-
- Lokeshwar V. B., Rubinowicz D., Schroeder G. L., Forgacs E., Minna J. D., Block N. L., Nadji M., Lokeshwar B. L. (2001) J. Biol. Chem. 276, 11922–11932 - PubMed
-
- Lipponen P., Aaltomaa S., Tammi R., Tammi M., Agren U., Kosma V. M. (2001) Eur. J. Cancer 37, 849–856 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
