

Protein kinase Czeta regulates Cdk5/p25 signaling during myogenesis
- PMID: 20200223
- PMCID: PMC2854099
- DOI: 10.1091/mbc.e09-10-0847
Protein kinase Czeta regulates Cdk5/p25 signaling during myogenesis
Abstract
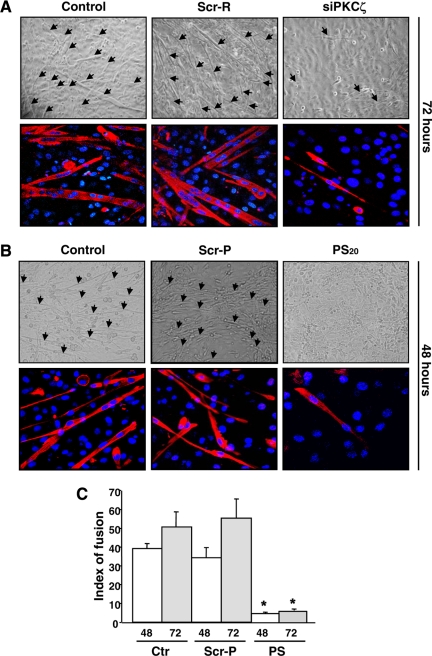
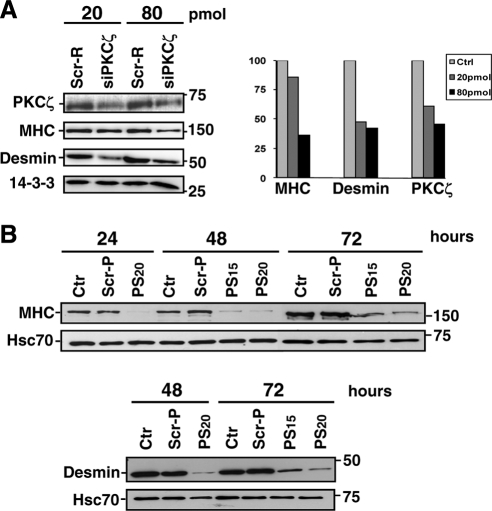
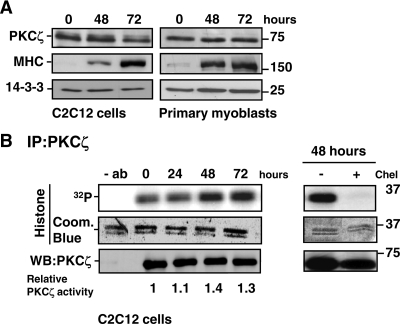
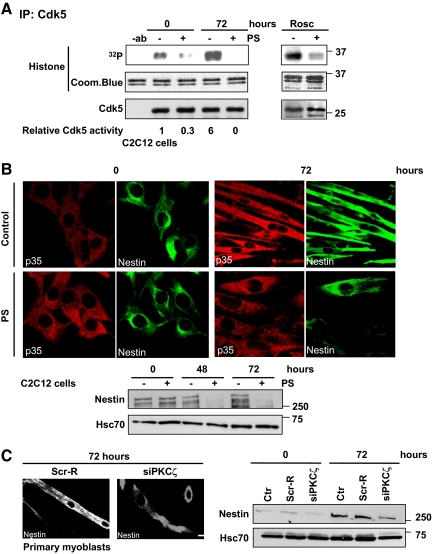
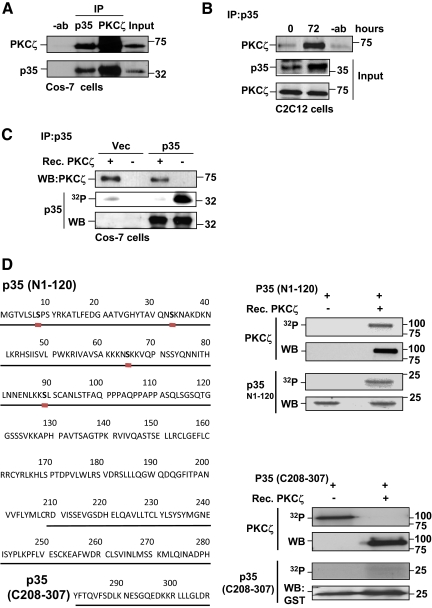
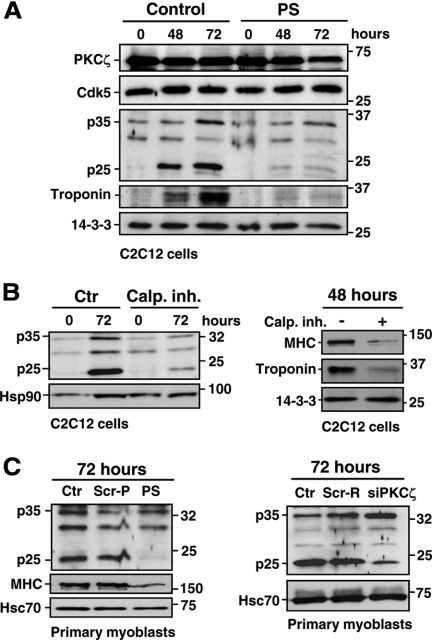
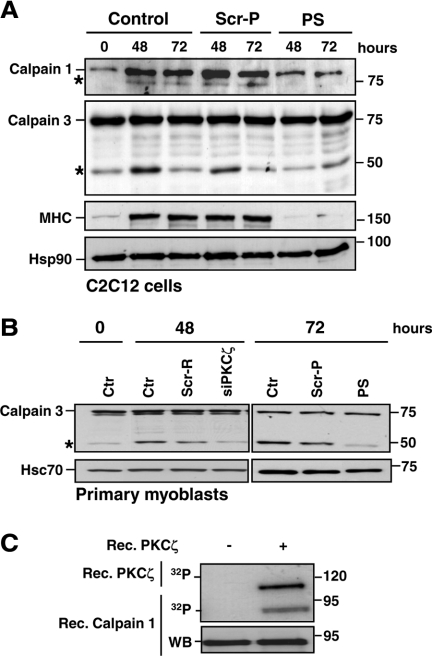
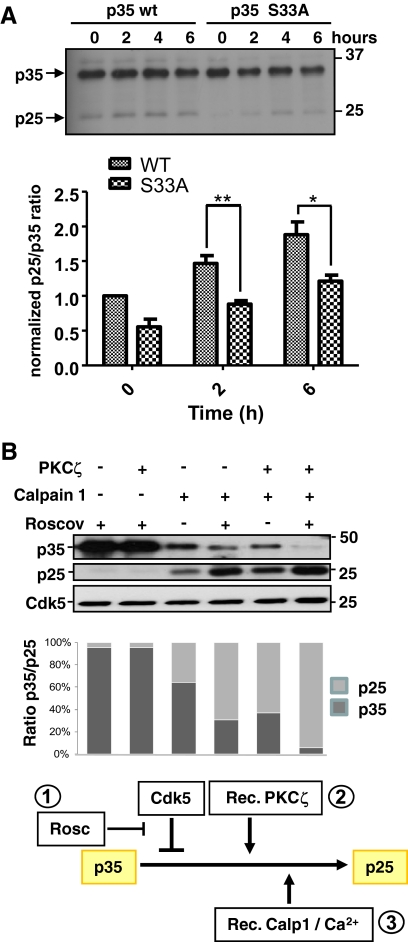
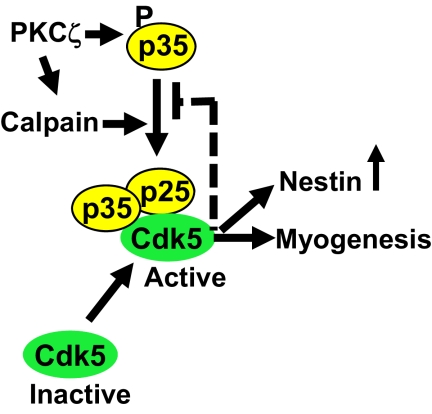
Atypical protein kinase Czeta (PKCzeta) is emerging as a mediator of differentiation. Here, we describe a novel role for PKCzeta in myogenic differentiation, demonstrating that PKCzeta activity is indispensable for differentiation of both C2C12 and mouse primary myoblasts. PKCzeta was found to be associated with and to regulate the Cdk5/p35 signaling complex, an essential factor for both neuronal and myogenic differentiation. Inhibition of PKCzeta activity prevented both myotube formation and simultaneous reorganization of the nestin intermediate filament cytoskeleton, which is known to be regulated by Cdk5 during myogenesis. p35, the Cdk5 activator, was shown to be a specific phosphorylation target of PKCzeta. PKCzeta-mediated phosphorylation of Ser-33 on p35 promoted calpain-mediated cleavage of p35 to its more active and stable fragment, p25. Strikingly, both calpain activation and the calpain-mediated cleavage of p35 were shown to be PKCzeta-dependent in differentiating myoblasts. Overall, our results identify PKCzeta as a controller of myogenic differentiation by its regulation of the phosphorylation-dependent and calpain-mediated p35 cleavage, which is crucial for the amplification of the Cdk5 activity that is required during differentiation.
Figures
Similar articles
-
Nestin as a regulator of Cdk5 in differentiating myoblasts.Mol Biol Cell. 2011 May;22(9):1539-49. doi: 10.1091/mbc.E10-07-0568. Epub 2011 Feb 23. Mol Biol Cell. 2011. PMID: 21346193 Free PMC article.
-
Cdk5 regulates the organization of Nestin and its association with p35.Mol Cell Biol. 2003 Jul;23(14):5090-106. doi: 10.1128/MCB.23.14.5090-5106.2003. Mol Cell Biol. 2003. PMID: 12832492 Free PMC article.
-
The regulation of cyclin-dependent kinase 5 activity through the metabolism of p35 or p39 Cdk5 activator.Neurosignals. 2003 Sep-Oct;12(4-5):221-9. doi: 10.1159/000074624. Neurosignals. 2003. PMID: 14673209 Review.
-
The chemokine SDF1 controls multiple steps of myogenesis through atypical PKCzeta.J Cell Sci. 2007 Nov 15;120(Pt 22):4050-9. doi: 10.1242/jcs.010009. Epub 2007 Oct 30. J Cell Sci. 2007. PMID: 17971416
-
The role of CDK5/P25 formation/inhibition in neurodegeneration.Drug News Perspect. 2006 Oct;19(8):453-60. doi: 10.1358/dnp.2006.19.8.1043961. Drug News Perspect. 2006. PMID: 17160145 Review.
Cited by
-
The myogenic kinome: protein kinases critical to mammalian skeletal myogenesis.Skelet Muscle. 2011 Sep 8;1:29. doi: 10.1186/2044-5040-1-29. Skelet Muscle. 2011. PMID: 21902831 Free PMC article.
-
CAPNS1 regulates USP1 stability and maintenance of genome integrity.Mol Cell Biol. 2013 Jun;33(12):2485-96. doi: 10.1128/MCB.01406-12. Epub 2013 Apr 15. Mol Cell Biol. 2013. PMID: 23589330 Free PMC article.
-
Intermediate Filaments and the Regulation of Cell Motility during Regeneration and Wound Healing.Cold Spring Harb Perspect Biol. 2017 Sep 1;9(9):a022046. doi: 10.1101/cshperspect.a022046. Cold Spring Harb Perspect Biol. 2017. PMID: 28864602 Free PMC article. Review.
-
A kinase of many talents: non-neuronal functions of CDK5 in development and disease.Open Biol. 2020 Jan;10(1):190287. doi: 10.1098/rsob.190287. Epub 2020 Jan 8. Open Biol. 2020. PMID: 31910742 Free PMC article. Review.
-
Lung omics signatures in a bronchopulmonary dysplasia and pulmonary hypertension-like murine model.Am J Physiol Lung Cell Mol Physiol. 2018 Nov 1;315(5):L734-L741. doi: 10.1152/ajplung.00183.2018. Epub 2018 Jul 26. Am J Physiol Lung Cell Mol Physiol. 2018. PMID: 30047283 Free PMC article.
References
-
- Barnoy S., Glaser T., Kosower N. S. The calpain-calpastatin system and protein degradation in fusing myoblasts. Biochim. Biophys. Acta. 1998;1402:52–60. - PubMed
-
- Barnoy S., Maki M., Kosower N. S. Overexpression of calpastatin inhibits L8 myoblast fusion. Biochem. Biophys. Res. Commun. 2005;332:697–701. - PubMed
-
- Berra E., Diaz-Meco M. T., Dominguez I., Municio M. M., Sanz L., Lozano J., Chapkin R. S., Moscat J. Protein kinase C zeta isoform is critical for mitogenic signal transduction. Cell. 1993;74:555–563. - PubMed
-
- Castellani L., Salvati E., Alema S., Falcone G. Fine regulation of RhoA and Rock is required for skeletal muscle differentiation. J. Biol. Chem. 2006;281:15249–15257. - PubMed
-
- Cheung Z. H., Ip N. Y. Cdk5, mediator of neuronal death and survival. Neurosci. Lett. 2004;361:47–51. [Review] - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
