

rBMP represses Wnt signaling and influences skeletal progenitor cell fate specification during bone repair
- PMID: 20200943
- PMCID: PMC3153130
- DOI: 10.1002/jbmr.29
rBMP represses Wnt signaling and influences skeletal progenitor cell fate specification during bone repair
Abstract
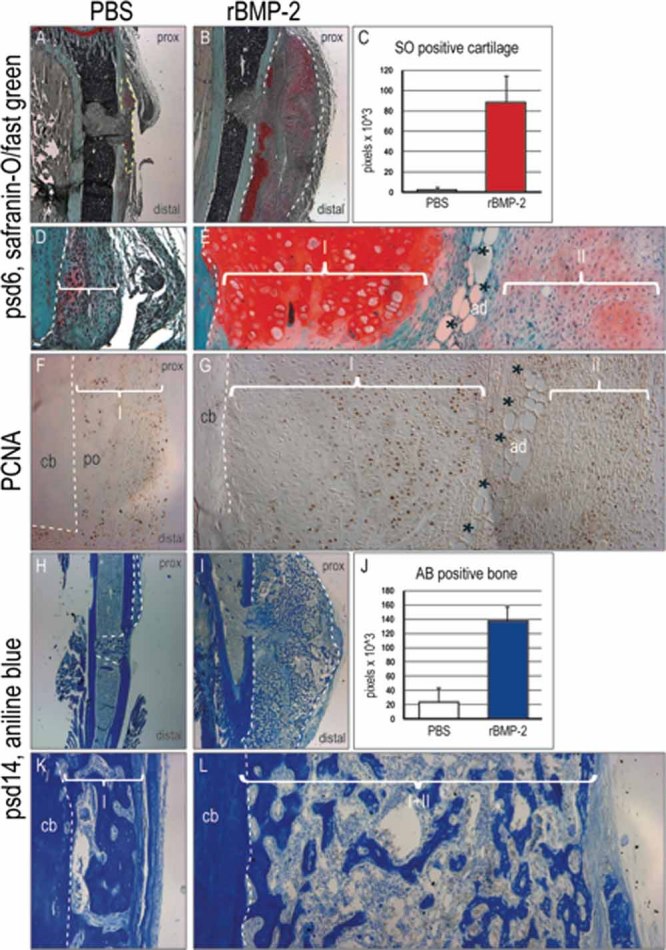
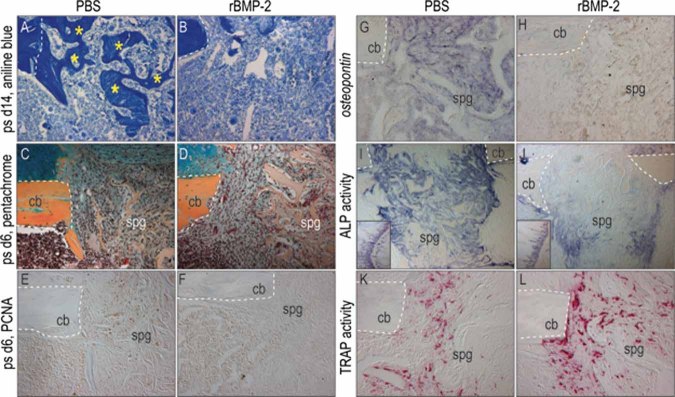
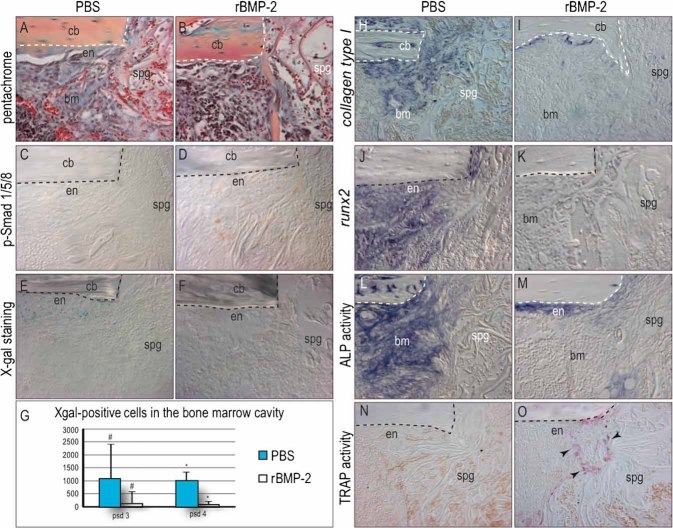
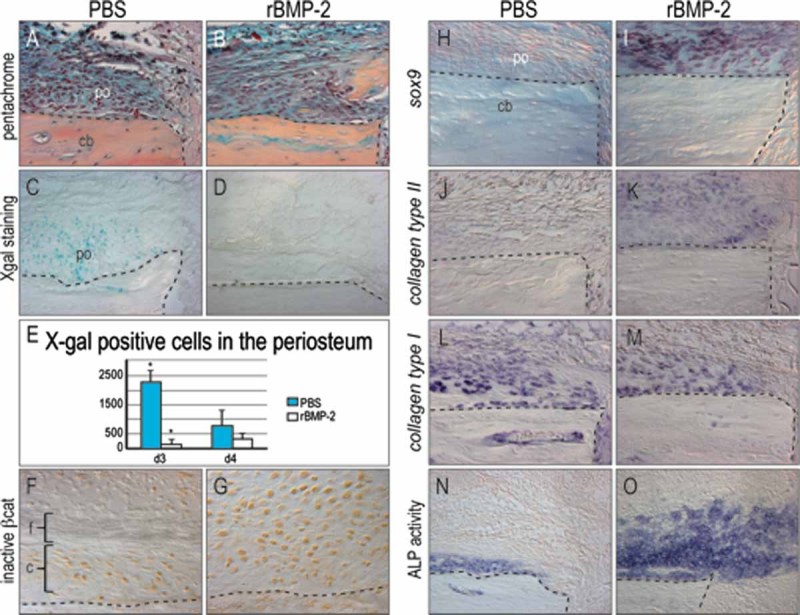
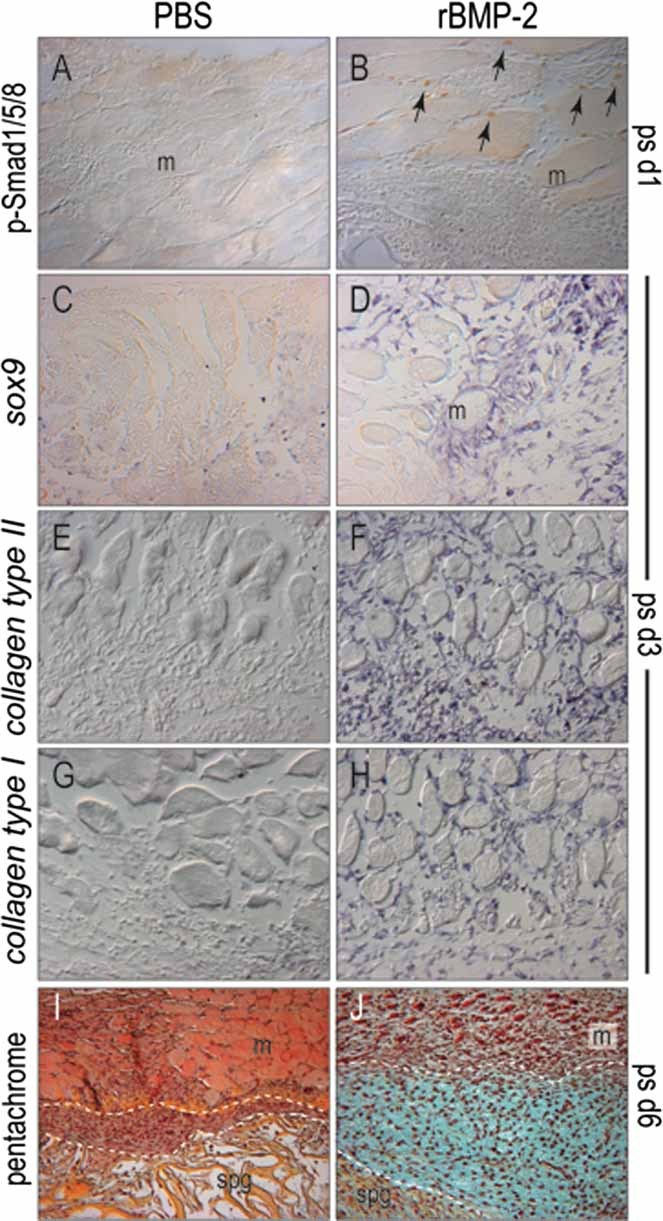
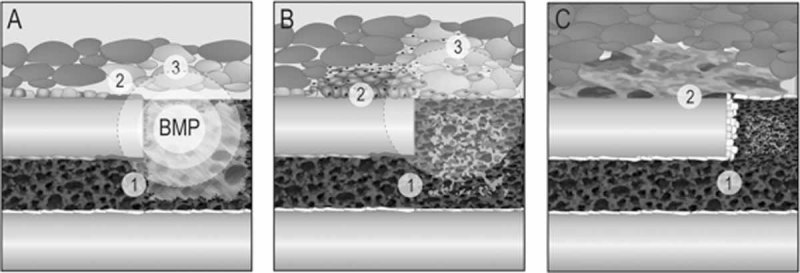
Bone morphogenetic proteins (BMPs) participate in multiple stages of the fetal skeletogenic program from promoting cell condensation to regulating chondrogenesis and bone formation through endochondral ossification. Here, we show that these pleiotropic functions are recapitulated when recombinant BMPs are used to augment skeletal tissue repair. In addition to their well-documented ability to stimulate chondrogenesis in a skeletal injury, we show that recombinant BMPs (rBMPs) simultaneously suppress the differentiation of skeletal progenitor cells in the endosteum and bone marrow cavity to an osteoblast lineage. Both the prochondrogenic and antiosteogenic effects are achieved because rBMP inhibits endogenous beta-catenin-dependent Wnt signaling. In the injured periosteum, this repression of Wnt activity results in sox9 upregulation; consequently, cells in the injured periosteum adopt a chondrogenic fate. In the injured endosteum, rBMP also inhibits Wnt signaling, which results in the runx2 and collagen type I downregulation; consequently, cells in this region fail to differentiate into osteoblasts. In muscle surrounding the skeletal injury site, rBMP treatment induces Smad phosphorylation followed by exuberant cell proliferation, an increase in alkaline phosphatase activity, and chondrogenic differentiation. Thus different populations of adult skeletal progenitor cells interpret the same rBMP stimulus in unique ways, and these responses mirror the pleiotropic effects of BMPs during fetal skeletogenesis. These mechanistic insights may be particularly useful for optimizing the reparative potential of rBMPs while simultaneously minimizing their adverse outcomes.
(c) 2010 American Society for Bone and Mineral Research.
Figures
Comment in
-
Novel complexities regarding BMPs and fracture healing.J Bone Miner Res. 2010 Jun;25(6):1193-5. doi: 10.1002/jbmr.122. J Bone Miner Res. 2010. PMID: 20499366 No abstract available.
References
-
- Ducy P, Zhang R, Geoffroy V, Ridall AL, Karsenty G. Osf2/Cbfa1: a transcriptional activator of osteoblast differentiation. Cell. 1997;89:747–754. - PubMed
-
- Komori T, Yagi H, Nomura S, et al. Targeted disruption of Cbfa1 results in a complete lack of bone formation owing to maturational arrest of osteoblasts. Cell. 1997;89:755–764. - PubMed
-
- Vortkamp A, Lee K, Lanske B, Segre GV, Kronenberg HM, Tabin CJ. Regulation of rate of cartilage differentiation by Indian hedgehog and PTH-related protein. Science. 1996;273:613–622. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
